Comparisons of the Postprandial Inflammatory and Endotoxaemic Responses to Mixed Meals in Young and Older Individuals: A Randomised Trial

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
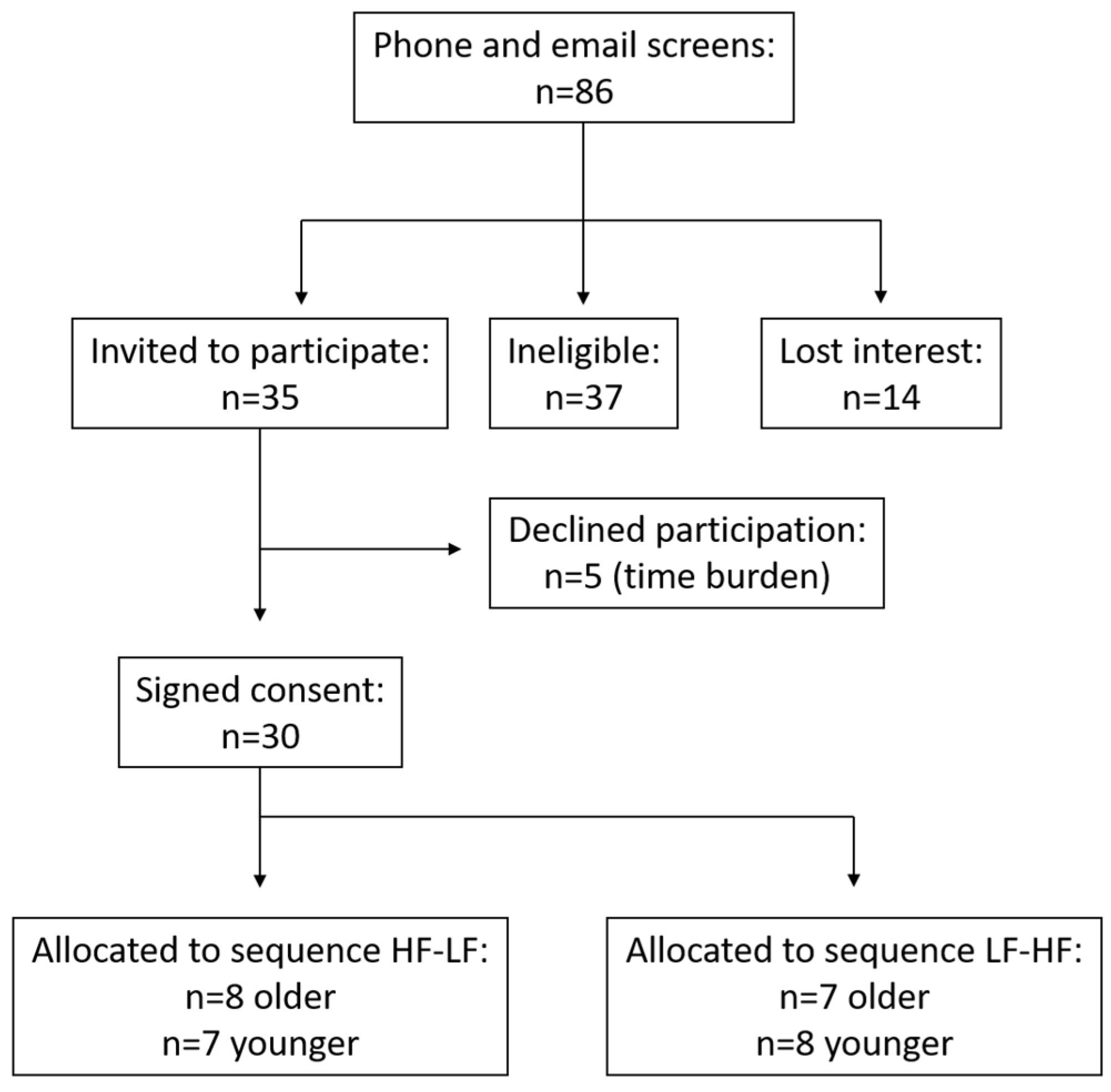
2.1. Subject Selection
2.2. Study Design and Treatments
2.3. Study Procedures
2.4. Chylomicron Isolation
2.5. PBMC Isolation and RNA Extraction
2.6. Quantitative Real-Time Reverse-Transcriptase Polymerase Chain Reaction (qPCR) Analysis
2.7. Biochemical Analysis
2.8. Endotoxin Analysis
2.9. Statistical Analyses
3. Results
3.1. Subject Characteristics
3.2. Postprandial Endotoxaemia
3.3. PBMC Senescence RNA Expression
3.4. PBMC RNA Expression of Endotoxaemic Activation
3.5. PBMC RNA Expression of Activation by Lipoproteins
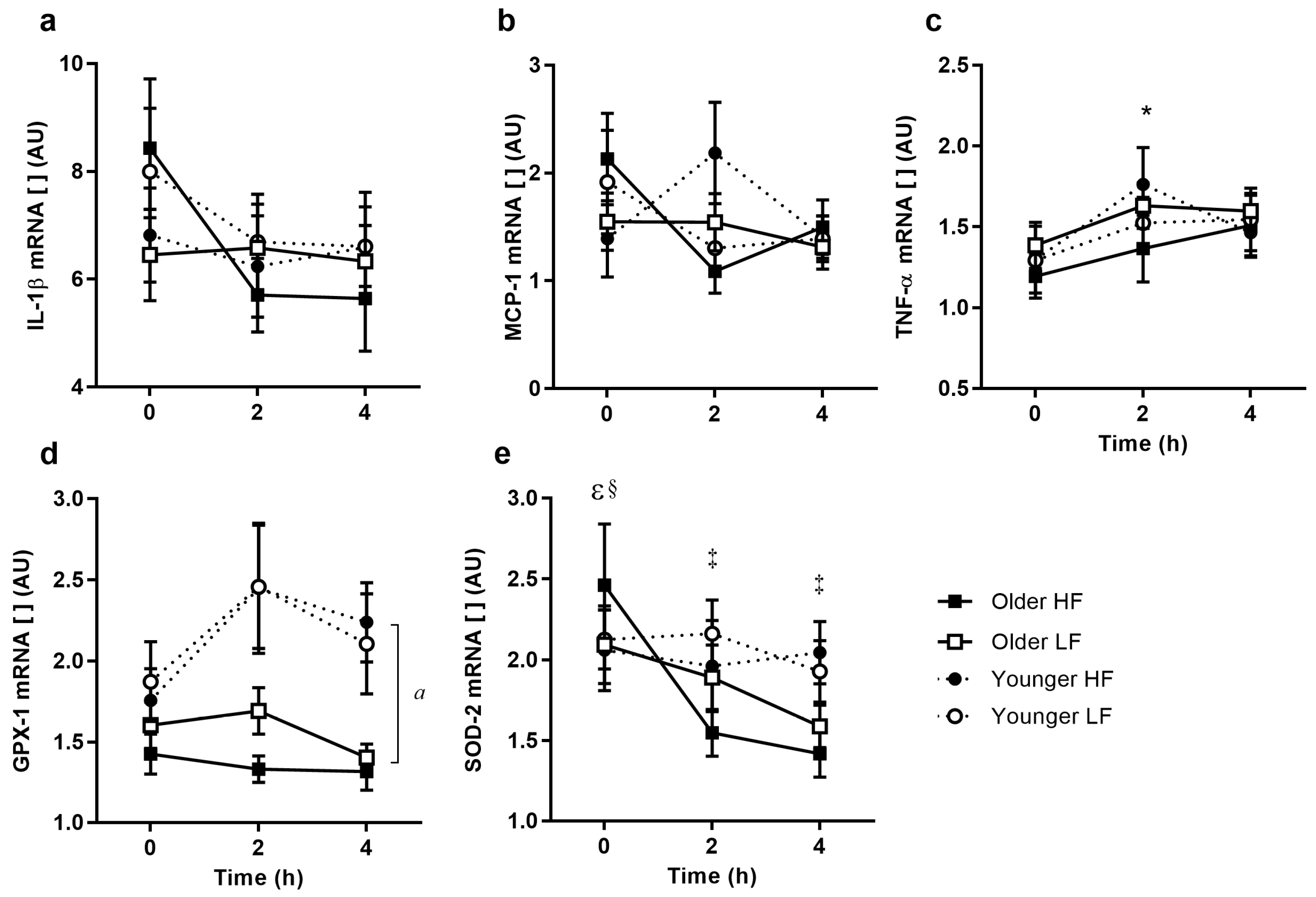
3.6. PBMC Inflammatory Cytokine Gene Expression
3.7. PBMC RNA Expression of Oxidative Stress Response
3.8. Cytokine and Antioxidant Protein Expression
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Krabbe, K.S.; Pedersen, M.; Bruunsgaard, H. Inflammatory mediators in the elderly. Exp. Gerontol. 2004, 39, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Bruunsgaard, H.; Pedersen, A.N.; Schroll, M.; Skinhøj, P.; Pedersen, B.K. Impaired production of proinflammatory cytokines in response to lipopolysaccharide (LPS) stimulation in elderly humans. Clin. Exp. Immunol. 1999, 118, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Duncan, B.B.; Schmidt, M.I.; Pankow, J.S.; Ballantyne, C.M.; Couper, D.; Vigo, A.; Hoogeveen, R.; Folsom, A.R.; Heiss, G. Low-grade systemic inflammation and the development of type 2 diabetes: The atherosclerosis risk in communities study. Diabetes 2003, 52, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Samdal Steinskog, E.S.; Wiig, H. Adipose tissue macrophages: The inflammatory link between obesity and cancer? Expert Opin. Ther. Targets 2015, 19, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Akbaraly, T.N.; Hamer, M.; Ferrie, J.E.; Lowe, G.; Batty, G.D.; Hagger-Johnson, G.; Singh-Manoux, A.; Shipley, M.J.; Kivimäki, M. Chronic inflammation as a determinant of future aging phenotypes. Can. Med. Assoc. J. 2013, 185, E763–E770. [Google Scholar] [CrossRef] [PubMed]
- Cannizzo, E.S.; Clement, C.C.; Sahu, R.; Follo, C.; Santambrogio, L. Oxidative stress, inflamm-aging and immunosenescence. J. Proteom. 2011, 74, 2313–2323. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Newman, A.B. Inflammatory markers in population studies of aging. Ageing Res. Rev. 2011, 10, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.G.; Myers, A.J.; Magnusson, P.K.E.; Prince, J.A. Transcriptome-wide assessment of human brain and lymphocyte senescence. PLoS ONE 2008, 3, E3024. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rahman, A.; Abdul Karim, N.; Abdul Hamid, N.A.; Harun, R.; Wan Ngah, W.Z. Senescence-related changes in gene expression of peripheral blood mononuclear cells from octo/nonagenarians compared to their offspring. Oxid. Med. Cell. Longev. 2013, 2013, 189129. [Google Scholar] [CrossRef] [PubMed]
- Candore, G.; Colonna-Romano, G.; Balistreri, C.R.; Di Carlo, D.; Grimaldi, M.P.; Listì, F.; Nuzzo, D.; Vasto, S.; Lio, D.; Caruso, C. Biology of longevity: Role of the innate immune system. Rejuvenation Res. 2006, 9, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Proctor, S.D.; Mamo, J.C.; Botham, K.M.; Cabezas, M.C. Understanding postprandial inflammation and its relationship to lifestyle behaviour and metabolic diseases. Int. J. Vasc. Med. 2012, 2012, 947417. [Google Scholar] [CrossRef] [PubMed]
- Laugerette, F.; Vors, C.; Peretti, N.; Michalski, M.-C. Complex links between dietary lipids, endogenous endotoxins and metabolic inflammation. Biochimie 2011, 93, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, 5–78. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, S.; Mensink, M.; Esser, D.; Feskens, E.; Muller, M.; Afman, L.A. Responses to high-fat challenges varying in fat type in subjects with different metabolic risk phenotypes: A randomized trial. PLoS ONE 2012, 7, E41388. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A. Effects of macronutrient excess and composition on oxidative stress: Relevance to diabetes and cardiovascular disease. Curr. Atheroscler Rep. 2006, 8, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Manning, P.J.; Sutherland, W.H.F.; McGrath, M.M.; De Jong, S.A.; Walker, R.J.; Williams, M.J.A. Postprandial cytokine concentrations and meal composition in obese and lean women. Obesity 2008, 16, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Patel, C.; Ghanim, H.; Ravishankar, S.; Chang, L.S.; Viswanathan, P.; Mohanty, P.; Dandona, P. Prolonged reactive oxygen species generation and nuclear factor-κb activation after a high-fat, high-carbohydrate meal in the obese. J. Clin. Endocrinol. Metab. 2007, 92, 4476–4479. [Google Scholar] [CrossRef] [PubMed]
- Stolk, R.P.; Pols, H.A.P.; Lamberts, S.W.J.; De Jong, P.T.V.M.; Hofman, A.; Grobbee, D.E. Diabetes mellitus, impaired glucose tolerance, and hyperinsulinemia in an elderly population: The rotterdam study. Am. J. Epidemiol. 1997, 145, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Marques-Vidal, P.; Mazoyer, E.; Bongard, V.; Gourdy, P.; Ruidavets, J.B.; Drouet, L.; Ferrières, J. Prevalence of insulin resistance syndrome in southwestern france and its relationship with inflammatory and hemostatic markers. Diabetes Care 2002, 25, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Laugerette, F.; Vors, C.; Géloën, A.; Chauvin, M.-A.; Soulage, C.; Lambert-Porcheron, S.; Peretti, N.; Alligier, M.; Burcelin, R.; Laville, M.; et al. Emulsified lipids increase endotoxemia: Possible role in early postprandial low-grade inflammation. J. Nutr. Biochem. 2011, 22, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Levels, J.H.; Abraham, P.R.; van den Ende, A.; van Deventer, S.J. Distribution and kinetics of lipoprotein-bound endotoxin. Infect. Immun. 2001, 69, 2821–2828. [Google Scholar] [CrossRef] [PubMed]
- Deopurkar, R.; Ghanim, H.; Friedman, J.; Abuaysheh, S.; Sia, C.L.; Mohanty, P.; Viswanathan, P.; Chaudhuri, A.; Dandona, P. Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of toll-like receptor-4 and suppressor of cytokine signaling-3. Diabetes Care 2010, 33, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Harte, A.L.; Varma, M.C.; Tripathi, G.; McGee, K.C.; Al-Daghri, N.M.; Al-Attas, O.S.; Sabico, S.; O’Hare, J.P.; Ceriello, A.; Saravanan, P.; et al. High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care 2012, 35, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Mani, V.; Hollis, J.H.; Gabler, N.K. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Nutr. Metab. 2013, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Grassi, M.; Petraccia, L.; Mennuni, G.; Fontana, M.; Scarno, A.; Sabetta, S.; Fraioli, A. Changes, functional disorders, and diseases in the gastrointestinal tract of elderly. Nutr. Hosp. 2011, 26, 659–668. [Google Scholar] [PubMed]
- Woudstra, T.; Thomson, A.B.R. Nutrient absorption and intestinal adaptation with ageing. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saffrey, M.J. Aging of the mammalian gastrointestinal tract: A complex organ system. Age 2014, 36, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.M. Changes in gastrointestinal function attributed to aging. Am. J. Clin. Nutr. 1992, 55, 1203S–1207S. [Google Scholar] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Pendyala, S.; Walker, J.M.; Holt, P.R. A high-fat diet is associated with endotoxemia that originates from the gut. Gastroenterology 2012, 142, 1100–1101.e2. [Google Scholar] [CrossRef] [PubMed]
- Cohn, J.S.; McNamara, J.R.; Cohn, S.D.; Ordovas, J.M.; Schaefer, E.J. Postprandial plasma lipoprotein changes in human subjects of different ages. J. Lipid Res. 1988, 29, 469–479. [Google Scholar] [PubMed]
- Puga, G.M.; Meyer, C.; Mandarino, L.J.; Katsanos, C.S. Increased plasma availability of l-arginine in the postprandial period decreases the postprandial lipemia in older adults. Nutrition 2013, 29, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Puga, G.M.; Meyer, C.; Everman, S.; Mandarino, L.J.; Katsanos, C.S. Postprandial lipemia in the elderly involves increased incorporation of ingested fat in plasma free fatty acids and small (Sf 20-400) triglyceride-rich lipoproteins. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E356–E361. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Mekki, N.; Boirie, Y.; Partier, A.; Grolier, P.; Alexandre-Gouabau, M.C.; Beaufrere, B.; Armand, M.; Lairon, D.; Azais-Braesco, V. Postprandial chylomicron and plasma vitamin e responses in healthy older subjects compared with younger ones. Eur. J. Clin. Investig. 1997, 27, 812–821. [Google Scholar] [CrossRef]
- Krasinski, S.D.; Cohn, J.S.; Schaefer, E.J.; Russell, R.M. Postprandial plasma retinyl ester response is greater in older subjects compared with younger subjects: Evidence for delayed plasma clearance of intestinal lipoproteins. J. Clin. Investig. 1990, 85, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Mekki, N.; Boirie, Y.; Partier, A.; Alexandre-Gouabau, M.C.; Grolier, P.; Beaufrere, B.; Portugal, H.; Lairon, D.; Azais-Braesco, V. Comparison of the postprandial plasma vitamin a response in young and older adults. J. Gerontol. A Biol. Sci. Med. Sci. 1998, 53, B133–B140. [Google Scholar] [CrossRef] [PubMed]
- Cardinault, N.; Tyssandier, V.; Grolier, P.; Winklhofer-Roob, B.M.; Ribalta, J.; Bouteloup-Demange, C.; Rock, E.; Borel, P. Comparison of the postprandial chylomicron carotenoid responses in young and older subjects. Eur. J. Nutr. 2003, 42, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Haahr, M. Random integer set generator. Available online: www.random.org/integer-sets/ (accessed on 22 August 2012).
- Milan, A.M.; Nuora, A.; Pundir, S.; Pileggi, C.A.; Markworth, J.F.; Linderborg, K.M.; Cameron-Smith, D. Older adults have an altered chylomicron response to a high-fat meal. Br. J. Nutr. 2016, 115, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Abuaysheh, S.; Sia, C.L.; Korzeniewski, K.; Chaudhuri, A.; Fernandez-Real, J.M.; Dandona, P. Increase in plasma endotoxin concentrations and the expression of toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal: Implications for insulin resistance. Diabetes Care 2009, 32, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Milan, A.M.; D’Souza, R.F.; Pundir, S.; Pileggi, C.A.; Barnett, M.P.G.; Markworth, J.F.; Cameron-Smith, D.; Mitchell, C. Older adults have delayed amino acid absorption after a high protein mixed breakfast meal. J. Nutr. Health Aging 2015, 19, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Mizunuma, Y.; Iwasaki, Y.; Tharwat, M. Changes of very low-density lipoprotein concentration in hepatic blood from cows with fasting-induced hepatic lipidosis. Can. J. Vet. Res. 2010, 74, 317–320. [Google Scholar] [PubMed]
- Naito, H.K. Lipoprotein Separations Using the TL-100 Tabletop Ultrafuge; Beckman Coulter: Pasadena, CA, USA, 1986. [Google Scholar]
- Kupke, I.R.; Wörz-Zeugner, S. Sequential microultracentrifugation of lipoproteins in 100ul of serum. J. Lipid Res. 1986, 27, 988–995. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-δδCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ketchum, P.A.; Novitsky, T.J. Assay of endotoxin by limulus amebocyte lysate. In Septic Shock Methods and Protocols; Thomas, J.E., Ed.; Humana Press: New York, NY, USA, 2000; Volume 36, pp. 3–12. [Google Scholar]
- Matthews, D.; Hosker, J.; Rudenski, A.; Naylor, B.; Treacher, D.; Turner, R. Homeostasis model assessment: Insulin resistance and b-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.G.; Poppitt, S.D.; Minihane, A.M. Postprandial lipemia and cardiovascular disease risk: Interrelationships between dietary, physiological and genetic determinants. Atherosclerosis 2012, 220, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Raz, O.; Steinvil, A.; Berliner, S.; Tovit, R.; Justo, D.; Shapira, I. The effect of two iso-caloric meals containing equal amounts of fats with a different fat composition on the inflammatory and metabolic markers in apparently healthy volunteers. J. Inflamm. 2013, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.; Peña-Orihuela, P.; Rangel-Zúñiga, O.A.; Pérez-Martínez, P.; Delgado-Lista, J.; Cruz-Teno, C.; Marín, C.; Tinahones, F.; Malagón, M.M.; Roche, H.M.; et al. Peripheral blood mononuclear cells as in vivo model for dietary intervention induced systemic oxidative stress. Food Chem. Toxicol. 2014, 72, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Burton-Freeman, B.; Talbot, J.; Park, E.; Krishnankutty, S.; Edirisinghe, I. Protective activity of processed tomato products on postprandial oxidation and inflammation: A clinical trial in healthy weight men and women. Mol. Nutr. Food Res. 2012, 56, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Schwander, F.; Kopf-Bolanz, K.A.; Buri, C.; Portmann, R.; Egger, L.; Chollet, M.; McTernan, P.G.; Piya, M.K.; Gijs, M.A.; Vionnet, N. A dose-response strategy reveals differences between normal-weight and obese men in their metabolic and inflammatory responses to a high-fat meal. J. Nutr. 2014, 144, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.A.; Klop, B.; Janssen, H.W.; Njo, T.L.; Westerman, E.M.; Castro Cabezas, M. Postprandial inflammation: Targeting glucose and lipids. Adv. Exp. Med. Biol. 2014, 824, 161–170. [Google Scholar] [PubMed]
- De Vries, M.A.; Klop, B.; Eskes, S.A.; van der Loos, T.L.J.M.; Klessens-Godfroy, F.J.M.; Wiebolt, J.; Janssen, H.W.; Westerman, E.M.; Castro Cabezas, M. The postprandial situation as a pro-inflammatory condition. Clin. Investig. Arterioscler. 2014, 26, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Alipour, A.; Van Oostrom, A.J.H.H.M.; Izraeljan, A.; Verseyden, C.; Collins, J.M.; Frayn, K.N.; Plokker, T.W.M.; Elte, J.W.F.; Cabezas, M.C. Leukocyte activation by triglyceride-rich lipoproteins. Atertioscler. Thromb. Vasc. Biol. 2008, 28, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.M.; Da Silva, N.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [PubMed]
- Erridge, C.; Attina, T.; Spickett, C.M.; Webb, D.J. A high-fat meal induces low-grade endotoxemia: Evidence of a novel mechanism of postprandial inflammation. Am. J. Clin. Nutr. 2007, 86, 1286–1292. [Google Scholar] [PubMed]
- Katz, D.; Hollander, D.; Said, H.M.; Dadufalza, V. Aging-associated increase in intestinal permeability to polyethylene glycol 900. Dig. Dis. Sci. 1987, 32, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Greenwood-Van Meerveld, B. Age-associated remodeling of the intestinal epithelial barrier. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Schröder, N.W.J.; Morath, S.; Alexander, C.; Hamann, L.; Hartung, T.; Zähringer, U.; Göbel, U.B.; Weber, J.R.; Schumann, R.R. Lipoteichoic acid (LTA) of streptococcus pneumoniae and staphylococcus aureus activates immune cells via toll-like receptor (TLR)-2, lipopolysaccharide-binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J. Biol. Chem. 2003, 278, 15587–15594. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Quintela, A.; Alonso, M.; Campos, J.; Vizcaino, L.; Loidi, L.; Gude, F. Determinants of serum concentrations of lipopolysaccharide-binding protein (LBP) in the adult population: The role of obesity. PLoS ONE 2013, 8, E54600. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, S.P.; Wen, T.; Zhou, H.N.; Hu, M.; Li, J.X. Postprandial hypertriglyceridemia associated with inflammatory response and procoagulant state after a high-fat meal in hypertensive patients. Coron. Artery Dis. 2008, 19, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, S.; Wang-Polagruto, J.; Polagruto, J.; Keen, C.L.; Jialal, I. High-fat, energy-dense, fast-food-style breakfast results in an increase in oxidative stress in metabolic syndrome. Metabolism 2008, 57, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef] [PubMed]
- Yubero-Serrano, E.M.; Gonzalez-Guardia, L.; Rangel-Zuñiga, O.; Delgado-Casado, N.; Delgado-Lista, J.; Perez-Martinez, P.; Garcia-Rios, A.; Caballero, J.; Marin, C.; Gutierrez-Mariscal, F.M.; et al. Postprandial antioxidant gene expression is modified by mediterranean diet supplemented with coenzyme q10 in elderly men and women. Age 2013, 35, 159–170. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item Name | Weight (g) | Macronutrients (g) 1 | Energy (kcal) | ||
|---|---|---|---|---|---|
| Carbohydrates | Fat | Protein | |||
| High fat breakfast | |||||
| Sausage and Egg Muffin Sandwich (×2) | 162 | 25.2 | 21 | 23.4 | 390 |
| Hash Brown (×2) | 56 | 13.5 | 10.1 | 1.5 | 150 |
| Total | 77.4 | 62.2 | 49.8 | 1080 | |
| Low fat breakfast | |||||
| Rolled Oats | 37 | 20.8 | 1.9 | 5.0 | 120 |
| 1% Cottage Cheese | 167 | 4.5 | 1.0 | 19.7 | 110 |
| Mixed Grain Bread | 42 | 11.2 | 2.2 | 5.1 | 90 |
| Reduced Fat Peanut Butter, Smooth | 25 | 8.4 | 9.4 | 4.4 | 140 |
| Fresh Peach | 154 | 14.6 | 0.3 | 1.4 | 60 |
| Trim Milk | 365 | 17.9 | 1.8 | 14.2 | 150 |
| Total | 77.4 | 16.6 | 49.8 | 670 | |
| Measure 1 | Unit | Younger Subjects (n = 15) 2 | Older Subjects (n = 15) 2,3 |
|---|---|---|---|
| Age | years | 22.7 ± 0.4 | 67.3 ± 1.5 *** |
| BMI | kg/m2 | 23.7 ± 0.8 | 24.4 ± 1.0 |
| Glucose | mmol/L | 5.1 ± 0.1 | 5.2 ± 0.1 |
| HOMA-IR | 2.1 ± 0.2 | 1.9 ± 0.2 | |
| Cholesterol | mmol/L | 4.0 ± 0.1 | 5.0 ± 0.1 *** |
| LDL | mmol/L | 2.5 ± 0.1 | 3.0 ± 0.1 ** |
| HDL | mmol/L | 1.3 ± 0.0 | 1.8 ± 0.1 *** |
| TAG | mmol/L | 0.8 ± 0.0 | 0.9 ± 0.0 |
| Insulin | μU/mL | 9.2 ± 0.8 | 8.7 ± 1.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milan, A.M.; Pundir, S.; Pileggi, C.A.; Markworth, J.F.; Lewandowski, P.A.; Cameron-Smith, D. Comparisons of the Postprandial Inflammatory and Endotoxaemic Responses to Mixed Meals in Young and Older Individuals: A Randomised Trial. Nutrients 2017, 9, 354. https://doi.org/10.3390/nu9040354
Milan AM, Pundir S, Pileggi CA, Markworth JF, Lewandowski PA, Cameron-Smith D. Comparisons of the Postprandial Inflammatory and Endotoxaemic Responses to Mixed Meals in Young and Older Individuals: A Randomised Trial. Nutrients. 2017; 9(4):354. https://doi.org/10.3390/nu9040354
Chicago/Turabian StyleMilan, Amber M., Shikha Pundir, Chantal A. Pileggi, James F. Markworth, Paul A. Lewandowski, and David Cameron-Smith. 2017. "Comparisons of the Postprandial Inflammatory and Endotoxaemic Responses to Mixed Meals in Young and Older Individuals: A Randomised Trial" Nutrients 9, no. 4: 354. https://doi.org/10.3390/nu9040354