Effects of Food Components That Activate TRPA1 Receptors on Mucosal Ion Transport in the Mouse Intestine

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Ussing Chamber Experiments
2.3. Calcium Mobilisation Assay
2.4. Compounds
2.5. Data Analysis
3. Results
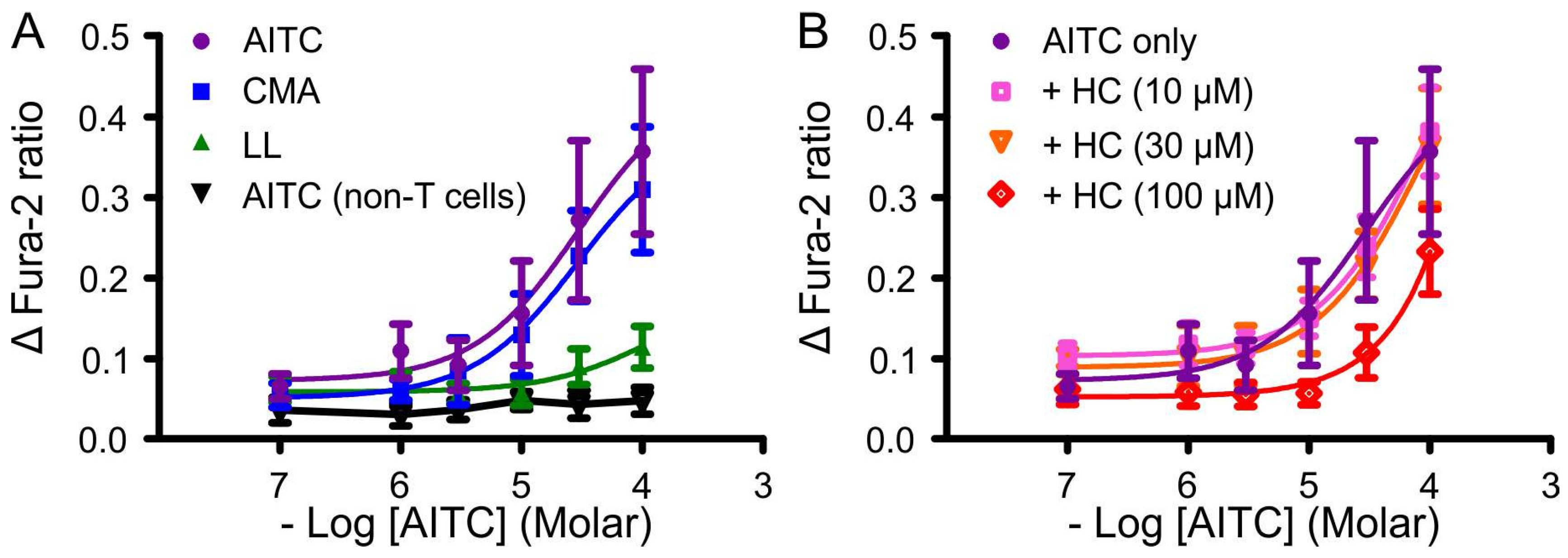
3.1. Effects of AITC, Cinnamaldehyde and Linalool on Ca2+ Mobilisation in HEK-TRPA1 Cells
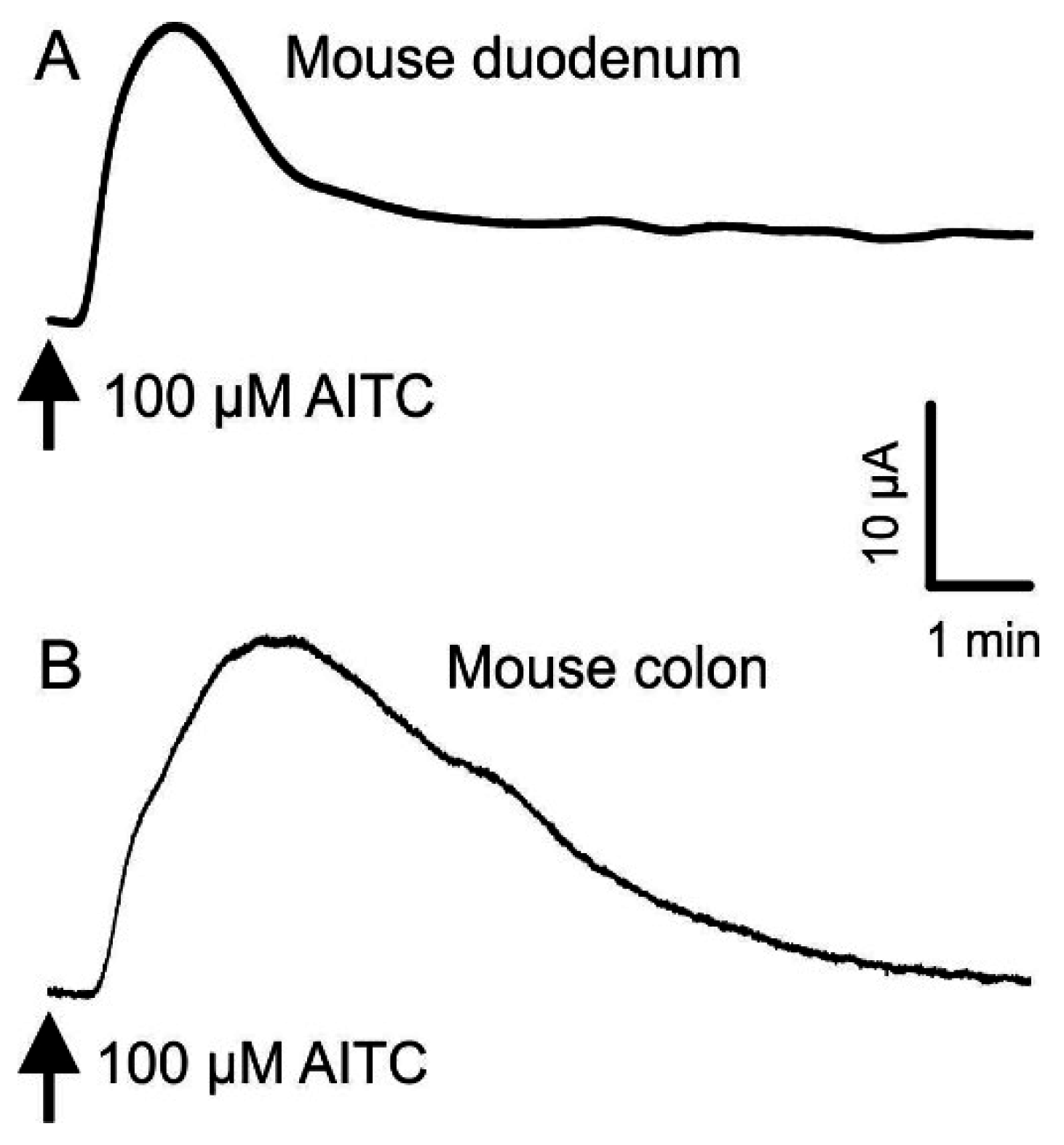
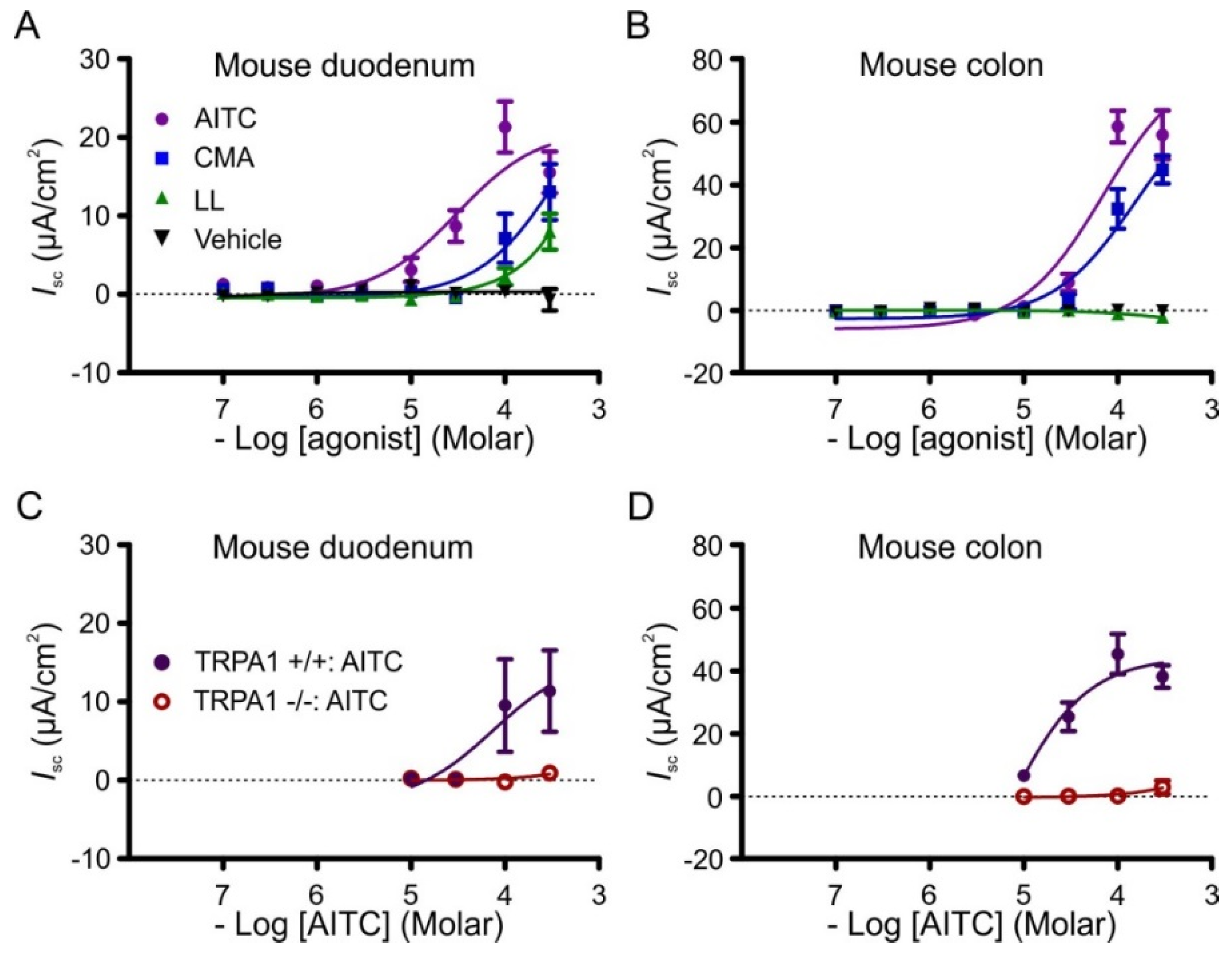
3.2. Effects of AITC, Cinnamaldehyde and Linalool on Short Circuit Current (Isc)
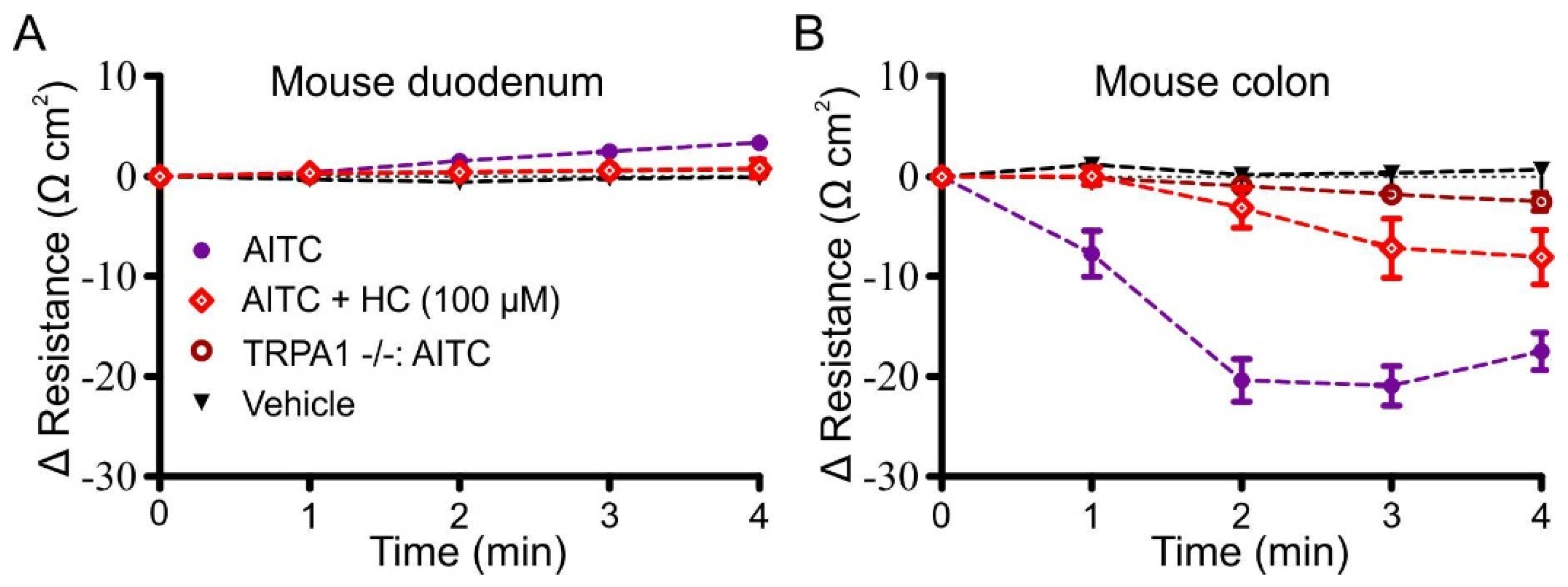
3.3. Effects of Pharmacological Antagonism and Ablation of Trpa1 on Responses to AITC
3.4. Effects of AITC, Cinnamaldehyde and Linalool on Transmucosal Resistance
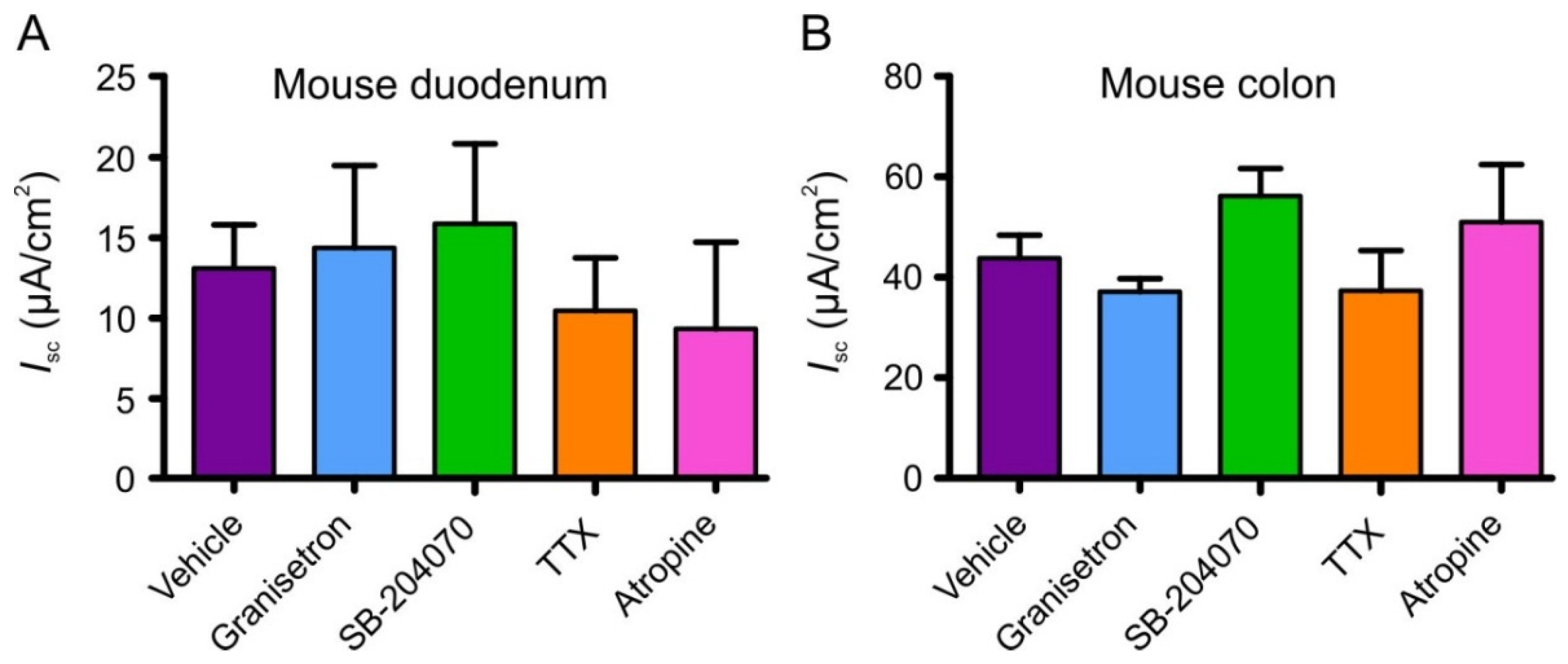
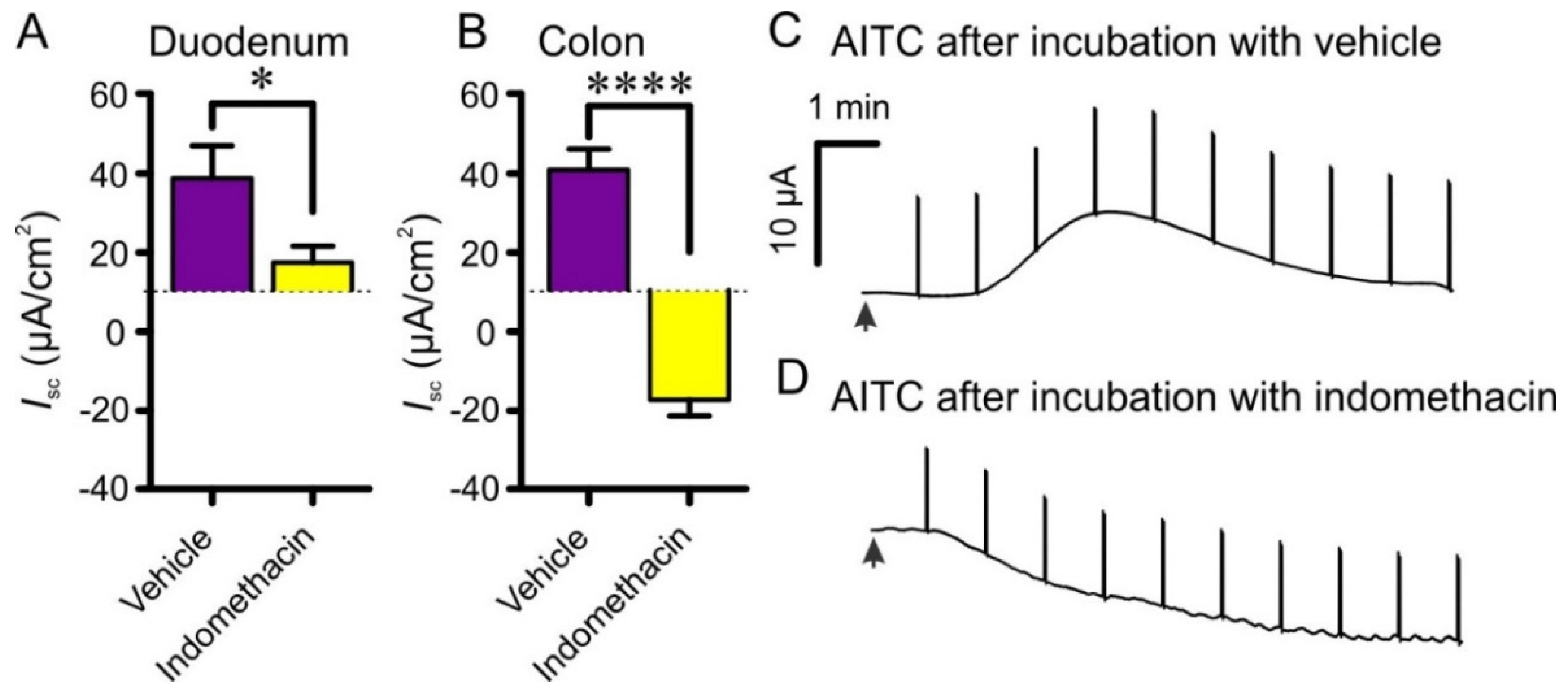
3.5. Effects of 5-HT Receptor Blockers, Tetrodotoxin, Atropine and Indomethacin
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86, E140–E148. [Google Scholar] [CrossRef] [PubMed]
- De Lange, C.F.M.; Pluske, J.; Gong, J.; Nyachoti, C.M. Strategic use of feed ingredients and feed additives to stimulate gut health and development in young pigs. Livest. Sci. 2010, 134, 124–134. [Google Scholar] [CrossRef]
- Hashemi, S.R.; Davoodi, H. Herbal plants and their derivatives as growth and health promoters in animal nutrition. Vet. Res. Commun. 2011, 35, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 report is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Holzer, P. TRP channels in the digestive system. Curr. Pharm. Biotechnol. 2011, 12, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Riera, C.E.; Menozzi-Smarrito, C.; Affolter, M.; Michlig, S.; Munari, C.; Robert, F.; Vogel, H.; Simon, S.A.; le Coutre, J. Compounds from Sichuan and Melegueta peppers activate, covalently and non-covalently, TRPA1 and TRPV1 channels. Br. J. Pharmacol. 2009, 157, 1398–1409. [Google Scholar] [CrossRef] [PubMed]
- Motter, A.L.; Ahern, G.P. TRPA1 is a polyunsaturated fatty acid sensor in mammals. PLoS ONE 2012, 7, e38439. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Fernandes, M.A.; Keeble, J.E. The functions of TRPA1 and TRPV1: Moving away from sensory nerves. Br. J. Pharmacol. 2012, 166, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Kaji, I.; Yasuoka, Y.; Karaki, S.I.; Kuwahara, A. Activation of TRPA1 by luminal stimuli induces EP4-mediated anion secretion in human and rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G690–G701. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.P.; Pelayo, J.C.; Cattaruzza, F.; Kuo, Y.M.; Gai, G.; Chiu, J.V.; Bron, R.; Furness, J.B.; Grady, E.F.; Bunnett, N.W. Transient receptor potential Ankyrin 1 is expressed by inhibitory motoneurons of the mouse intestine. Gastroenterology 2011, 141, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Callaghan, B.; Bron, R.; Bravo, D.M.; Furness, J.B. Identification of enteroendocrine cells that express TRPA1 channels in the mouse intestine. Cell Tissue Res. 2014, 356, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Purhonen, A.K.; Louhivuori, L.M.; Kiehne, K.; Akerman, K.E.O.; Herzig, K.H. TRPA1 channel activation induces cholecystokinin release via extracellular calcium. FEBS Lett. 2008, 582, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA 1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Perrier, C. Thyme and cinnamon extracts induce anion secretion in piglet small intestine via cholinergic pathways. J. Pharm. Pharmacol. 2008, 59, 543–552. [Google Scholar]
- Youn, H.S.; Lee, J.K.; Choi, Y.J.; Saitoh, S.I.; Miyake, K.; Hwang, D.H.; Lee, J.Y. Cinnamaldehyde suppresses toll-like receptor 4 activation mediated through the inhibition of receptor oligomerization. Biochem. Pharmacol. 2008, 75, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.P.; Amadesi, S.; Veldhuis, N.A.; Abogadie, F.C.; Lieu, T.M.; Darby, W.; Liedtke, W.; Lew, M.J.; McIntyre, P.; Bunnett, N.W. Protease-activated receptor 2 (PAR2) protein and transient receptor potential vanilloid 4 (TRPV4) protein coupling is required for sustained inflammatory signaling. J. Biol. Chem. 2013, 288, 5790–5802. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.; Hunne, B.; Hirayama, H.; Sartor, D.M.; Nguyen, T.V.; Abogadie, F.C.; Ferens, D.; McIntyre, P.; Ban, K.; Baell, J.; et al. Sites of action of ghrelin receptor ligands in cardiovascular control. Am. J. Physiol. 2012, 303, H1011–H1021. [Google Scholar] [CrossRef] [PubMed]
- Field, M. Intestinal ion transport and the pathophysiology of diarrhea. J. Clin. Invest. 2003, 111, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Wang, Y.Z.; Frieling, T.; Wood, J.D. Neural 5-hydroxytryptamine receptors regulate chloride secretion in guinea pig distal colon. Am. J. Physiol. 1991, 261, G833–G840. [Google Scholar] [PubMed]
- Neal, K.B.; Bornstein, J.C. Serotonergic receptors in therapeutic approaches to gastrointestinal disorders. Curr. Opin. Pharm. 2006, 6, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Cooke, H.J.; Su, H.C.; Fertel, R. Histamine augments colonic secretion in guinea pig distal colon. Am. J. Physiol. 1990, 258, G432–G439. [Google Scholar] [PubMed]
- Christofi, F.L.; Wunderlich, J.; Yu, J.G.; Wang, Y.Z.; Xue, J.; Guzman, J.; Javed, N.; Cooke, H. Mechanically evoked reflex electrogenic chloride secretion in rat distal colon is triggered by endogenous nucleotides acting at P2Y1, P2Y2, and P2Y4 receptors. J. Comp. Neurol. 2004, 469, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Staruschenko, A.; Jeske, N.A.; Akopian, N.C. Contribution of TRPV1-TRPA1 interaction to the single channel properties of the TRPA1 channel. J. Biol. Chem. 2010, 285, 15167–15177. [Google Scholar] [CrossRef] [PubMed]
- Mace, O.J.; Affleck, J.; Patel, N.; Kellett, G.L. Sweet taste receptors in rat small intestine stimulate glucose absorption through apical GLUT2. J. Physiol. (Lond.) 2007, 582, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Mace, O.J.; Morgan, E.L.; Affleck, J.A.; Lister, N.; Kellett, G.L. Calcium absorption by Cav1.3 induces terminal web myosinII phosphorylation and apical GLUT2 insertion in rat intestine. J. Physiol. (Lond.) 2007, 580, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.L.; Mace, O.J.; Affleck, J.; Kellett, G.L. Apical GLUT2 and Cav1.3: Regulation of rat intestinal glucose and calcium absorption. J. Physiol. (Lond.) 2007, 580, 593–604. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fothergill, L.J.; Callaghan, B.; Rivera, L.R.; Lieu, T.; Poole, D.P.; Cho, H.-J.; Bravo, D.M.; Furness, J.B. Effects of Food Components That Activate TRPA1 Receptors on Mucosal Ion Transport in the Mouse Intestine. Nutrients 2016, 8, 623. https://doi.org/10.3390/nu8100623
Fothergill LJ, Callaghan B, Rivera LR, Lieu T, Poole DP, Cho H-J, Bravo DM, Furness JB. Effects of Food Components That Activate TRPA1 Receptors on Mucosal Ion Transport in the Mouse Intestine. Nutrients. 2016; 8(10):623. https://doi.org/10.3390/nu8100623
Chicago/Turabian StyleFothergill, Linda J., Brid Callaghan, Leni R. Rivera, TinaMarie Lieu, Daniel P. Poole, Hyun-Jung Cho, David M. Bravo, and John B. Furness. 2016. "Effects of Food Components That Activate TRPA1 Receptors on Mucosal Ion Transport in the Mouse Intestine" Nutrients 8, no. 10: 623. https://doi.org/10.3390/nu8100623