Evaluation of Flavonoid Contents and Antioxidant Capacity of the Aerial Parts of Common and Tartary Buckwheat Plants

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Dry Weight (DW) Content
2.2. Total Flavonoid Content (TF)
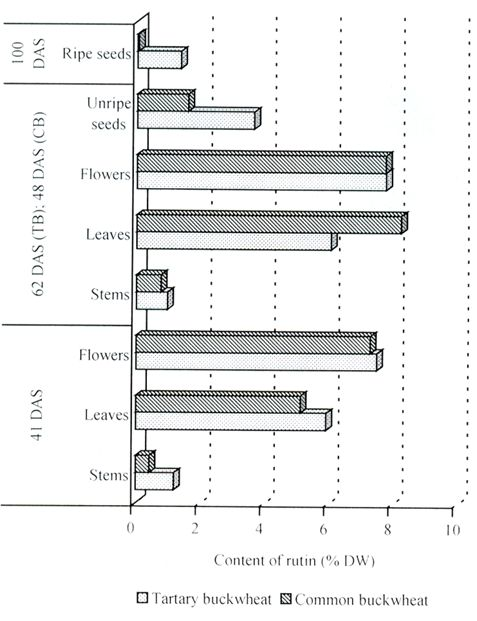
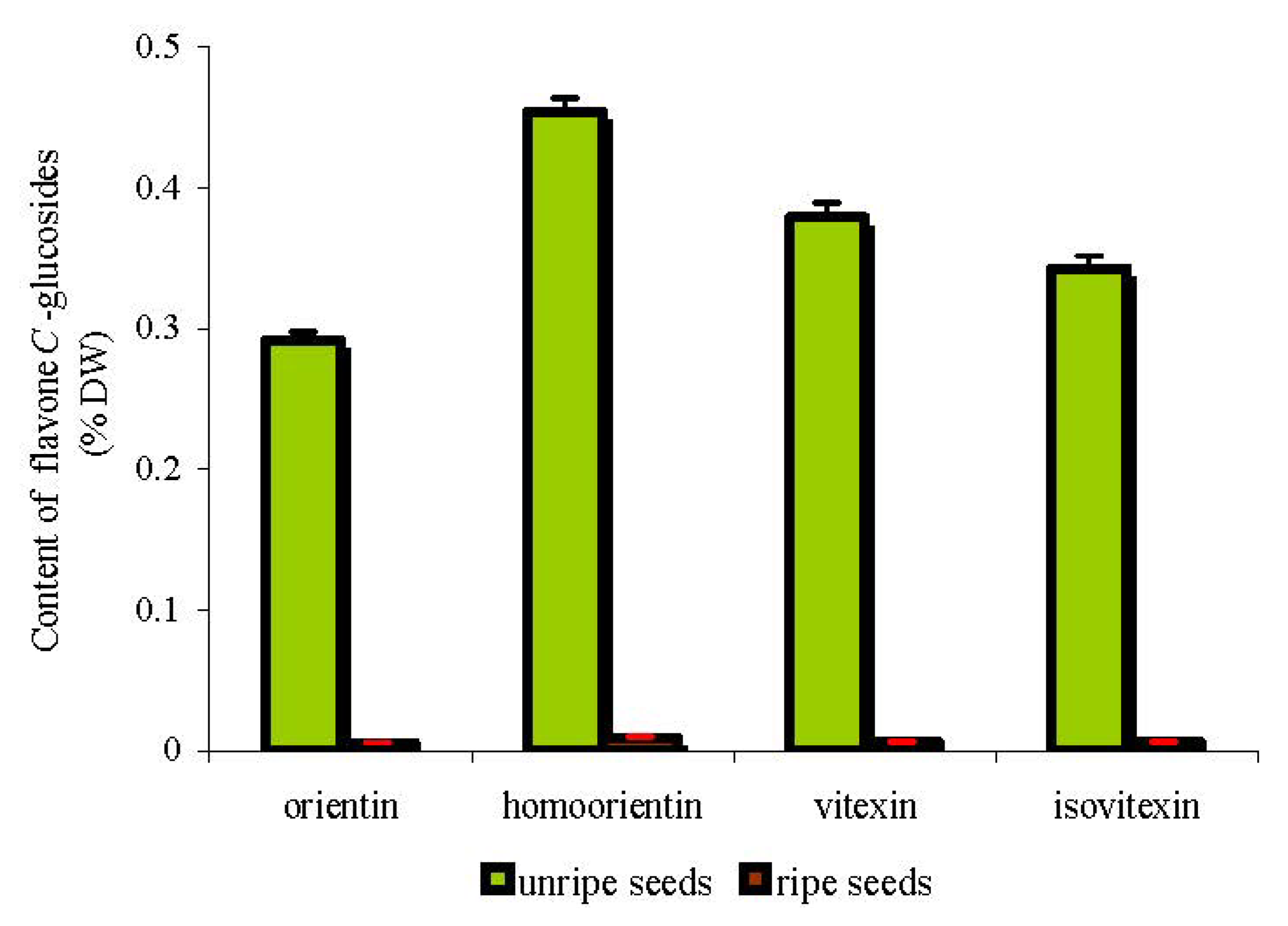
2.3. Rutin, Quercetin, Quercitrin and Flavone C-Glucosides Content
2.4. Antioxidant Capacity (AC) Determined by DPPH and PCL Assays
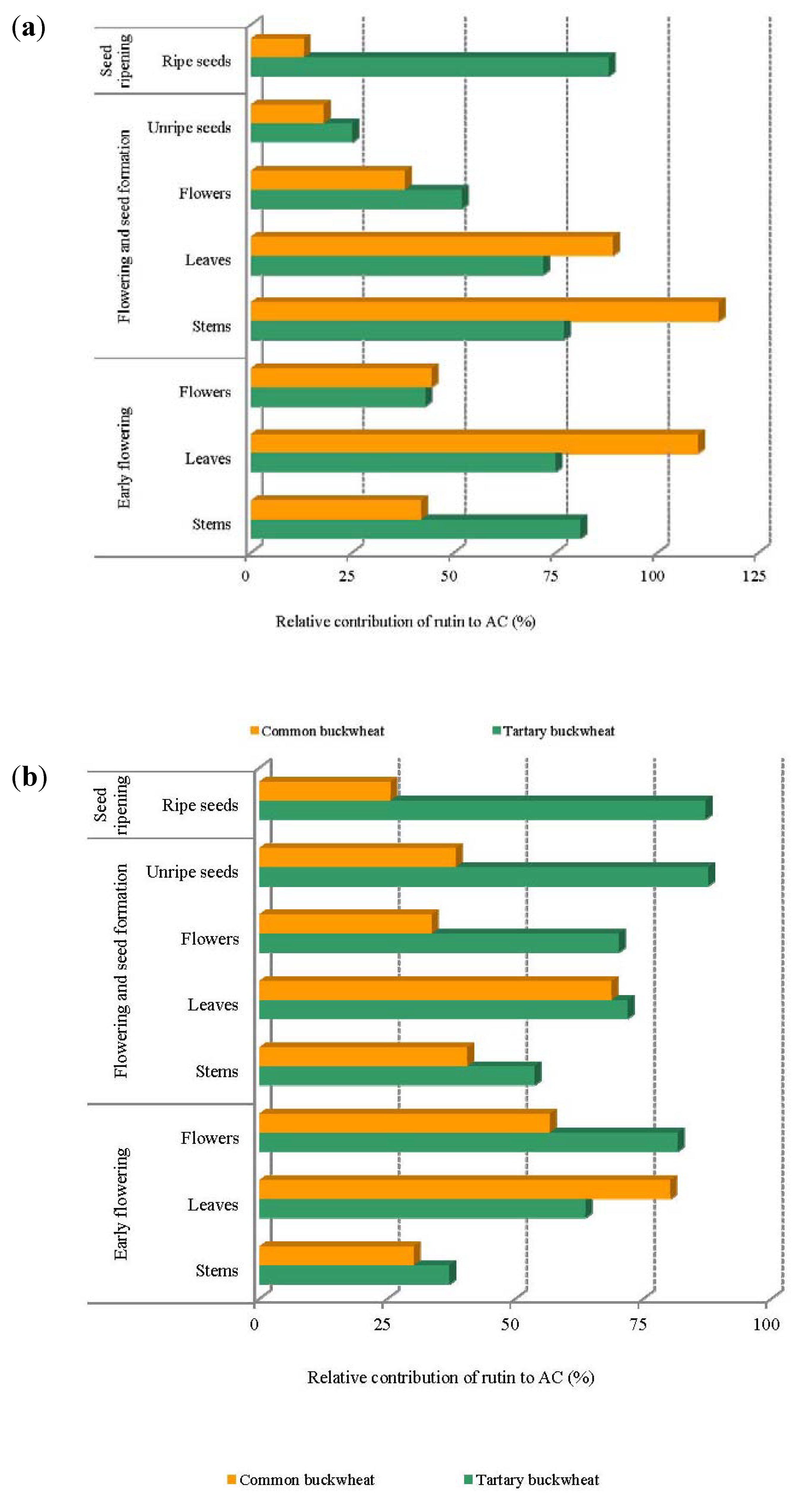
2.5. The Contribution of Rutin to the Antioxidant Capacity of Buckwheat Plant Material
3. Experimental
3.1. Chemicals
3.2. Buckwheat Plant Material
3.3. Extraction
3.4. Determination of Total Flavonoids (TF)
3.5. Quantitative Determination of Flavonoids by High-Performance Liquid Chromatography with Diode Array Detection (HPLC-DAD)
3.6. DPPH Radical Scavenging Assay
3.7. Photo-Induced Chemiluminescence Assay
3.8. Contribution of Rutin to the Antioxidant Capacity of Buckwheat Plant Material
3.9. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Przybylski, R.; Gruczyńska, E. A review of nutritional and nutraceutical components of buckwheat. Eur. J. Plant Sci. Biotechnol. 2009, 3, 10–22. [Google Scholar]
- Jiang, P.; Burczynski, F.; Campbell, C.; Pierce, G.; Austria, J.A.; Briggs, C.J. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F. tataricum, and F. homotropicum and their protective effects against lipid peroxidation. Food Res. Int. 2007, 40, 356–364. [Google Scholar] [CrossRef]
- Fabjan, N.; Rode, J.; Kosir, I.J.; Wang, Z.H.; Zhang, Z.; Kreft, A.I. Tartary buckwheat (Fagopyrum tataricum Gaertn.) as a source of dietary rutin and quercitrin. J. Agric. Food Chem. 2003, 51, 6452–6455. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Szawara-Nowak, D.; Zieliński, H. Determination of the antioxidant activity of rutin and its contribution to the antioxidant capacity of diversified buckwheat origin material by updated analytical strategies. Pol. J. Food Nutr. Sci. 2010, 60, 315–321. [Google Scholar]
- Zielińska, D.; Zieliński, H. Antioxidant activity of flavone C-glucosides determined by updated analytical strategies. Food Chem. 2011, 124, 672–678. [Google Scholar] [CrossRef]
- Afanas’ev, I.B.; Ostrakhovitch, E.A.; Mikhal’chik, E.V.; Ibragimova, G.A.; Korkina, G.L. Enhancement of antioxidant and anti-inflammatory activities of bioflavonoid rutin by complexation with transition metals. Biochem. Pharmacol. 2001, 61, 677–684. [Google Scholar] [CrossRef]
- Yoshiharu, M.; Hiroyasu, K.; Yoshitomi, I.; Tetsuo, M.; Kozo, O.; Hideo, M.; Katsumi, Y. Structure and hypotensive effect of flavonoids glycosides in Citrus unshiu peelings. Agric. Biol. Chem. 1985, 49, 909–914. [Google Scholar]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jia, J.; Yang, L.; Yang, F.; Ge, H.; Zhao, C.; Zhang, L.; Zu, Y. Evaluation of antioxidant activities of aqueous extracts and fractionation of different parts of Elsholtzia cilianta. Molecules 2012, 17, 5430–5441. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Chen, Y.S.; Yang, J.H.; Chiang, B.H. Antioxidant activity of tartary (Fagopyrum tataricum (L.) Gaertn.) and common (Fagopyrum esculentum Moench) buckwheat sprouts. J. Agric. Food Chem. 2008, 56, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ito, M. Effect of light on the content of phenolic compounds in buckwheat seedlings. Nippon Shokuhin Kagaku Kaishi 2003, 50, 23–34. [Google Scholar] [CrossRef]
- Kim, S.J.; Zaidul, I.S.M.; Suzuki, T.; Mukasa, Y.; Hashimoto, N.; Takigawa, S.; Noda, T.; Matsuura-Endo, C.; Yamauchi, H. Comparison of phenolic compositions between common and tartary buckwheat (Fagopyrum) sprouts. Food Chem. 2008, 110, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Watanabe, M.; Iki, M.; Aoyagi, Y.; Kim, S.J.; Mukasa, Y.; Kokota, S.; Takigawa, S.; Hashimoto, N.; Noda, T.; et al. Time-course study and effects of drying method on concentrations of γ-aminobutyric acid, flavonoids, anthocyanin, and 2"-hydroxynicotianamine in leaves of buckwheat. J. Agric. Food Chem. 2009, 57, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Bystrická, J.; Vollmannová, A.; Kupecsek, A.; Musilová, J.; Poláková, Z.; Čičová, I.; Bojňanska, T. Bioactive compounds in different plant parts of various buckwheat (Fagopyrum esculentum Moench.) cultivars. Cereal Res. Commun. 2011, 39, 436–444. [Google Scholar] [CrossRef]
- Holasová, M.; Fiedlerová, V.; Smrčinová, H.; Orsák, M.; Lachman, J.; Vavreinová, S. Buckwheat—The source of antioxidant activity in functional foods. Food Res. Int. 2002, 35, 207–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, J.; Liu, C.; Wu, X.; Zhang, Y. Isolation and purification of four flavone C-glycosides from antioxidant of bamboo leaves by macroporous resin column chromatography and preparative high-performance liquid chromatography. Food Chem. 2008, 107, 1326–1336. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Kalinová, J.; Vrchotová, N. Level of catechin, myricetin, quercetin and isoquercetin in buckwheat (Fagopyrum esculentum Moench), changes of their levels during vegetation and their effect on the growth of selected weeds. J. Agric. Food Chem. 2009, 57, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Szawara-Nowak, D.; Zieliński, H. Comparison of spectrophotometric and electrochemical methods for the evaluation of antioxidant capacity of buckwheat products after hydrothermal treatment. J. Agric. Food Chem. 2007, 55, 6124–6131. [Google Scholar] [CrossRef] [PubMed]
- Kreft, I.; Fabjan, N.; Yasumoto, K. Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 2006, 98, 508–512. [Google Scholar] [CrossRef]
- Kalinowá, J.; Dadáková, E. Varietal and year changes of rutin content in common buckwheat (Fagopyrum esculentum Moench). Cereal Res. Commun. 2006, 34, 1315–1321. [Google Scholar] [CrossRef]
- Brunori, A.; Sándor, G.; Xie, H.; Baviello, G.; Nehiba, B.; Rabnecz, G.; Végvári, G. Rutin content of the grain of 22 buckwheat (Fagopyrum esculentum Moench and Fagopyrum tataricum Gaertn.) varieties grown in Hungary. Eur. J. Plant Sci. Biotechnol. 2009, 3, 62–65. [Google Scholar]
- Cui, X.D.; Wang, Z.H. Preparation and properties of rutin-hydrolyzing enzyme from tartary buckwheat seeds. Food Chem. 2012, 132, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Honda, Y.; Funatsuki, W.; Nakatsuka, K. Purification and characterization of flavonol 3-glucosidase, and its activity during the ripening in tartary buckwheat seeds. Plant Sci. 2002, 163, 417–423. [Google Scholar] [CrossRef]
- Prabhakar, M.C.; Bamo, H.; Kumar, I.; Shamsi, M.A.; Khan, S.Y. Pharmacological investigations on vitexin. Planta Med. 1981, 43, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Agnese, A.M.; Perez, C.; Cabrera, J.L. Adesmia aegiceras antimicrobial activity and chemical study. Phytomedicine 2001, 8, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Hien, T.V.; Huong, N.B.; Hung, P.M.; Duc, N.B. Radioprotective effects of vitexina for breast cancer patients undergoing radioteraphy with cobalt-60. Integr. Cancer Ther. 2002, 1, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Jang, D.S.; Lee, Y.M.; Kim, J.M.; Kim, J.S. Naphthopyrone glucosides from the seeds of Cassia tora with inhibitory activity on advanced glycation end products (AGEs) formation. Arch. Pharm. Res. 2006, 29, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxides radicals. Food Chem. 1998, 64, 555–559. [Google Scholar]
- Popov, I.; Lewin, G. Antioxidative homeostasis: Characterization by means of chemiluminescent technique in methods in enzymology. In Oxidants and Antioxidants; Packer, L., Ed.; Academic Press: New York, NY, USA, 1999; pp. 96–100. [Google Scholar]
- Zielińska, D.; Szawara-Nowak, D.; Ornatowska, A.; Wiczkowski, W. The use of cyclic voltammetry, photochemiluminescence and spectrophotometric methods for the measurement of the antioxidant capacity of buckwheat sprouts. J. Agric. Food Chem. 2007, 55, 9891–9898. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Contact the authors. |

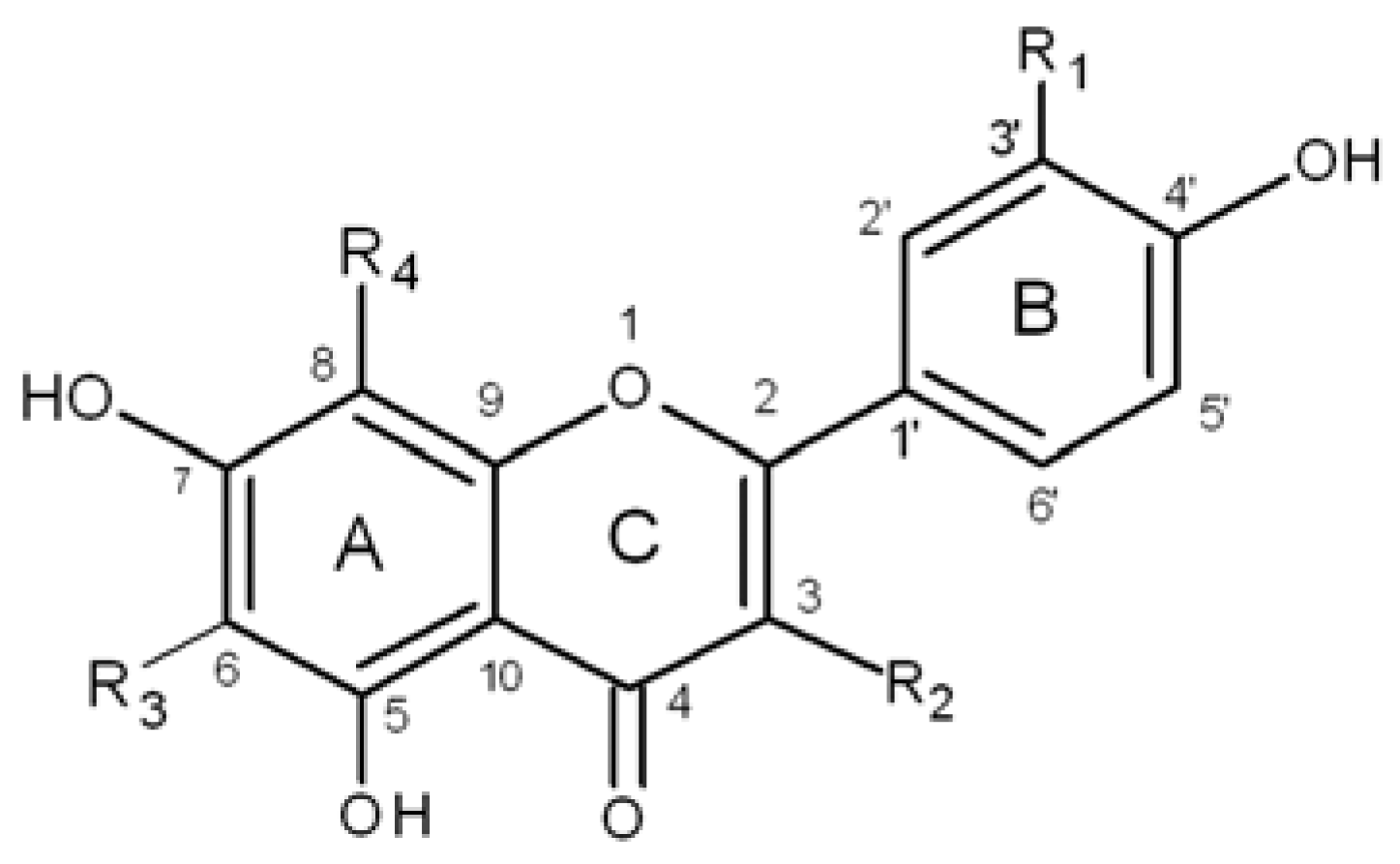
| Buckwheat Flavonoids | R1 | R2 | R3 | R4 |
| rutin | OH | rutinose | H | H |
| quercetin | OH | OH | H | H |
| quercitrin | OH | rhamnose | H | H |
| orientin | OH | H | H | glucose |
| homoorientin | OH | H | glucose | H |
| vitexin | H | H | H | glucose |
| isovitexin | H | H | glucose | H |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAS | Phenological State | Aerial Part | TF (mg RE/g DW) | |
|---|---|---|---|---|
| Common Buckwheat | Tartary Buckwheat | |||
| 41 (CB) 41 (TB) | early flowering | stems leaves flowers | 10.53 ± 1.03 aA 54.80 ± 2.54 bA 142.98 ± 5.81 dB | 17.63 ± 1.88 aB 81.08 ± 9.44 bB 131.32 ± 2.37 cA |
| 48 (CB) 62 (TB) | flowering and seed formation | stems leaves flowers unripe seeds | 8.82 ± 0.92 aA 81.89 ± 4.00 cA 203.63 ± 24.18 eA 47.80 ± 0.40 bA | 10.20 ± 1.55 aA 76.40 ± 16.10 bA 145.40 ± 28.75 cA 73.34 ± 1.15 bB |
| 100 (CB) 100 (TB) | seed ripening | ripe seeds | 5.78 ± 0.23 aA | 20.24 ± 0.31 aB |
| DAS | Phenological State | Aerial Part | Rutin (% DW) | |
|---|---|---|---|---|
| Common Buckwheat | Tartary Buckwheat | |||
| 41 (CB) 41 (TB) | early flowering | stems leaves flowers | 0.422 ± 0.048 aA 5.117 ± 0.461 cA 7.285 ± 0.248 dA | 1.183 ± 0.166 aB 5.924 ± 0.070 cB 7.485 ± 0.248 dA |
| 48 (CB) 62 (TB) | flowering and seed formation | stems leaves flowers unripe seeds | 0.763 ± 0.167 aA 8.237 ± 0.660 eB 7.761 ± 1.064 deA 1.573 ± 0.063 bA | 0.961 ± 0.014 aA 6.063 ± 0.832 cA 7.772 ± 0.993 dA 3.641 ± 0.202 bB |
| 100 (CB) 100 (TB) | seed ripening | ripe seeds | 0.043 ± 0.004 aA | 1.350 ± 0.087 aB |
| DAS | Phenological State | Aerial Part | AC (µmol TE/g DW) | |
|---|---|---|---|---|
| Common Buckwheat | Tartary Buckwheat | |||
| 41 (CB) 41 (TB) | early flowering | stems leaves flowers | 33.39 ± 8.16 aA 154.23 ± 8.33 bA 543.72 ± 5.38 dA | 48.62 ± 9.40 aA 262.71 ± 16.51 bB 577.05 ± 7.63 eB |
| 48 (CB) 62 (TB) | flowering and seed formation | stems leaves flowers unripe seeds | 21.95 ± 1.36 aA 307.08 ± 34.15 cA 675.77 ± 49.10 eB 285.68 ± 5.26 cA | 41.67 ± 4.93 aB 278.77 ± 40.51 bA 492.40 ± 30.82 dA 382.60 ± 4.10 cB |
| 100 (CB) 100 (TB) | seed ripening | ripe seeds | 11.70 ± 0.97 aA | 50.67 ± 2.89 aB |
| DAS | Phenological State | Aerial Part | AC (µmol TE/g DW) | |
|---|---|---|---|---|
| Common Buckwheat | Tartary Buckwheat | |||
| 41 (CB) 41 (TB) | early flowering | stems leaves flowers | 64.3 ± 8.8 abA 292.4 ± 66.6 cA 589.4 ± 3.2 dA | 146.4 ± 16.5 cB 426.9 ± 18.8 aB 420.1 ± 17.2 aB |
| 48 (CB) 62 (TB) | flowering and seed formation | stems leaves flowers unripe seeds | 86.3 ± 15.4 abA 549.4 ± 54.7 dA 1065.9 ± 212.9 eA 187.9 ± 9.2b cA | 82.1 ± 9.1 bA 387.0 ± 46.9 aB 508.3 ± 71.40 dB 190.4 ± 6.2 cA |
| 100 (CB) 100 (TB) | seed ripening | ripe seeds | 8.6 ± 0.2 aA | 71.1 ± 2.0 bB |
© 2012 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zielińska, D.; Turemko, M.; Kwiatkowski, J.; Zieliński, H. Evaluation of Flavonoid Contents and Antioxidant Capacity of the Aerial Parts of Common and Tartary Buckwheat Plants. Molecules 2012, 17, 9668-9682. https://doi.org/10.3390/molecules17089668
Zielińska D, Turemko M, Kwiatkowski J, Zieliński H. Evaluation of Flavonoid Contents and Antioxidant Capacity of the Aerial Parts of Common and Tartary Buckwheat Plants. Molecules. 2012; 17(8):9668-9682. https://doi.org/10.3390/molecules17089668
Chicago/Turabian StyleZielińska, Danuta, Marcin Turemko, Jacek Kwiatkowski, and Henryk Zieliński. 2012. "Evaluation of Flavonoid Contents and Antioxidant Capacity of the Aerial Parts of Common and Tartary Buckwheat Plants" Molecules 17, no. 8: 9668-9682. https://doi.org/10.3390/molecules17089668