
Neurological and Epigenetic Implications of Nutritional Deficiencies on Psychopathology: Conceptualization and Review of Evidence


{kind=link}
Abstract
:1. Introduction
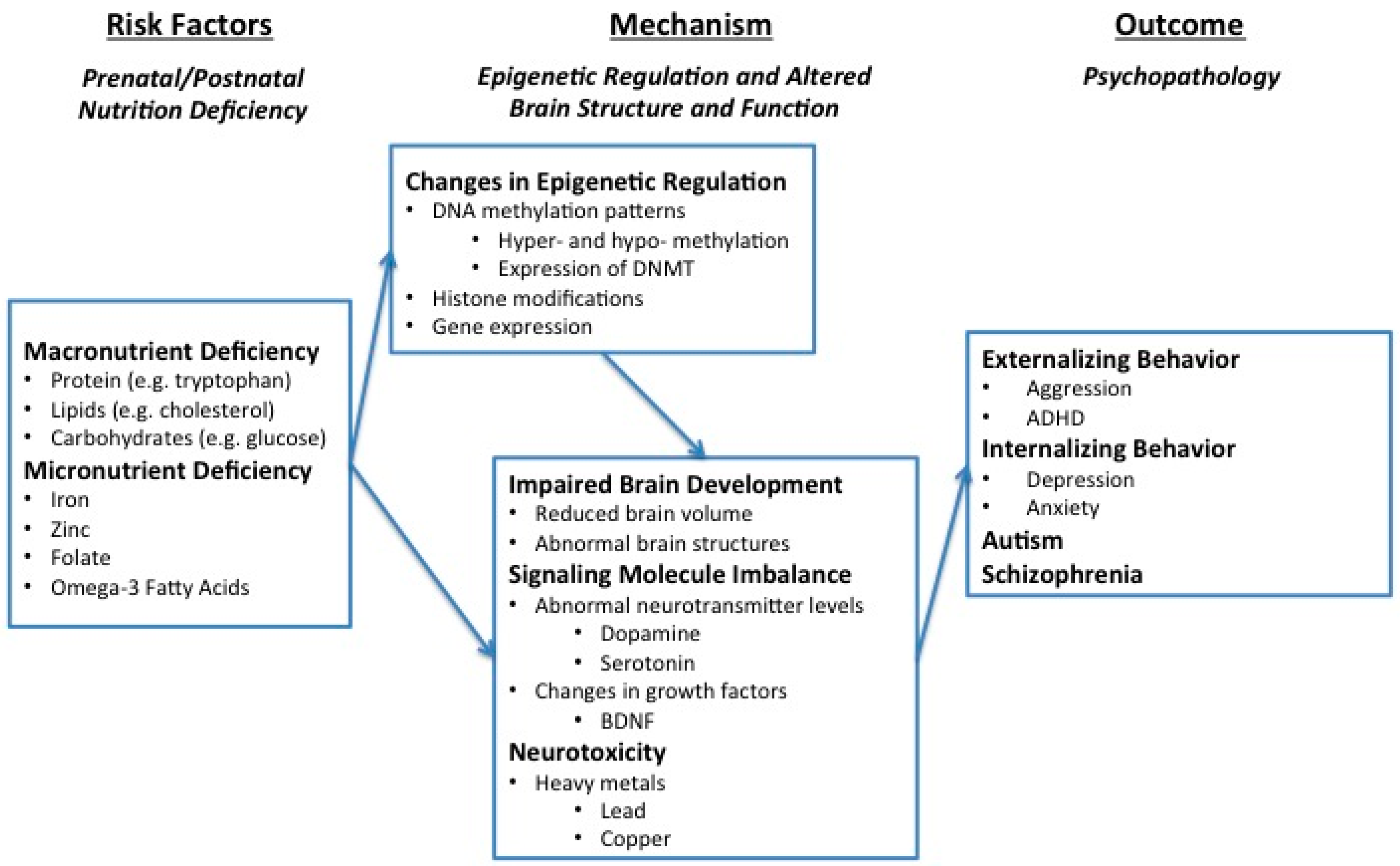
2. Overview of the Framework

3. Macro- and Micro-Nutrient Deficiency Are Risk Factors for Psychopathology
3.1. Macro-Nutrition Deficiency and Psychopathology
3.2. Micro-Nutrition Deficiency and Psychopathology
4. Mechanisms Mediating Nutrition Deficiency and Psychopathology: Epigenetics and Altered Brain Structure and Function
4.1. Nutrient Deficiency Alters Epigenetic Processes, Particularly in Early Development
4.1.1. Influence of Nutrition on DNA Methylation, Histone Modification, and Gene Expression
4.1.2. Influence of Epigenetic Changes on Brain Dysfunction and Psychopathology
4.2. Altered Brain Structure and Function as a Mediator
4.2.1. Impaired Brain Development
4.2.2. Signaling Molecule Imbalance
4.2.3. Increasing Neurotoxicity
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- De Castro, J.M. Genetic influences on daily intake and meal patterns of humans. Physiol. Behav. 1993, 53, 777–782. [Google Scholar] [CrossRef]
- Liu, J.; Tuvblad, C.; Raine, A.; Baker, L. Genetic and environmental influences on nutrient intake. Genes Nutr. 2013, 8, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A.; Raine, A. Biosocial bases of antisocial behavior: Psychophysiological, neurological, and cognitive factors. Clin. Psychol. Rev. 1997, 17, 589–604. [Google Scholar] [CrossRef]
- Rudo-Hutt, A.S.; Portnoy, J.; Chen, F.R.; Raine, A. Biosocial criminology as a paradigm shift. In The Routledge International Handbook of Biosocial Criminology; Routledge: New York, NY, USA, 2014; pp. 22–31. [Google Scholar]
- Portnoy, J.; Chen, F.R.; Raine, A. Biological protective factors for antisocial and criminal behavior. J. Crim. Justice 2013, 41, 292–299. [Google Scholar] [CrossRef]
- Liu, J. Early health risk factors for violence: Conceptualization, review of the evidence, and implications. Aggress. Violent Behav. 2011, 16, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Raine, A. The Anatomy of Violence: The Biological Roots of Crime; Random House, Inc.: New York, NY, USA, 2013. [Google Scholar]
- Liu, J.; Raine, A.; Venables, P.H.; Mednick, S.A. Malnutrition at age 3 years and externalizing behavior problems at ages 8, 11, and 17 years. Am. J. Psychiatry 2004, 161, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, R.; Hoek, H.W.; Susser, E. Prenatal exposure to wartime famine and development of antisocial personality disorder in early adulthood. JAMA 1999, 282, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Gesch, C.B.; Hammond, S.M.; Hampson, S.E.; Eves, A.; Crowder, M.J. Influence of supplementary vitamins, minerals and essential fatty acids on the antisocial behaviour of young adult prisoners. Randomised, placebo-controlled trial. Br. J. Psychiatry 2002, 181, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Raine, A. The effect of childhood malnutrition on externalizing behavior. Curr. Opin. Pediatr. 2006, 18, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Volavka, J.; Crowner, M.; Brizer, D.; Convit, A.; van Praag, H.; Suckow, R.F. Tryptophan treatment of aggressive psychiatric inpatients. Biol. Psychiatry 1990, 28, 728–732. [Google Scholar] [CrossRef]
- Naninck, E.F.; Lucassen, P.J.; Korosi, A. 15 Consequences of early-life experiences on cognition and emotion: A role for nutrition and epigenetic mechanisms. In The Oxford Handbook of Molecular Psychology; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Gallagher, E.A.; Newman, J.P.; Green, L.R.; Hanson, M.A. The effect of low protein diet in pregnancy on the development of brain metabolism in rat offspring. J. Physiol. 2005, 568, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Buydens-Branchey, L.; Branchey, M.; McMakin, D.L.; Hibbeln, J.R. Polyunsaturated fatty acid status and aggression in cocaine addicts. Drug Alcohol. Depend. 2003, 71, 319–323. [Google Scholar] [CrossRef]
- Rubia, K.; Lee, F.; Cleare, A.J.; Tunstall, N.; Fu, C.H.; Brammer, M.; McGuire, P. Tryptophan depletion reduces right inferior prefrontal activation during response inhibition in fast, event-related fMRI. Psychopharmacology (Berl.) 2005, 179, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Beers, M.; Berkow, R. The Merck Manual of Diagnosis and Therapy, Whitehouse Station, NJ; Merck and Co.: Rahway, NJ, USA, 1999. [Google Scholar]
- Neugebauer, R. Fetal Origins of Antisocial Personality Disorder and Schizophrenia: Evidence from the Dutch Hunger Winter 1944–45; Nova Publishers: New York, NY, USA, 2006. [Google Scholar]
- Shargel, L.; Wu-Pong, S.; Yu, A.B. Applied Biopharmaceutics & Pharmacokinetics; McGraw-Hill: New York, NY, USA, 2007. [Google Scholar]
- Hallberg, L.; Brune, M.; Rossander, L. Iron absorption in man: Ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr. 1989, 49, 140–144. [Google Scholar] [PubMed]
- Lopez, M.; Tena-Sempere, M. Estrogens and the control of energy homeostasis: A brain perspective. Trends Endocrinol. Metab. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tikal, K.; Benesova, O.; Frankova, S. The effect of pyrithioxine and pyridoxine on individual behavior, social interactions, and learning in rats malnourished in early postnatal life. Psychopharmacologia 1976, 46, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Duran, P.; Cintra, L.; Galler, J.R.; Tonkiss, J. Prenatal protein malnutrition induces a phase shift advance of the spontaneous locomotor rhythm and alters the rest/activity ratio in adult rats. Nutr. Neurosci. 2005, 8, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Khanna, V.K.; Husain, R.; Seth, P.K. Effect of protein malnutrition on the neurobehavioural toxicity of styrene in young rats. J. Appl. Toxicol. 1994, 14, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Alamy, M.; Bengelloun, W.A. Malnutrition and brain development: An analysis of the effects of inadequate diet during different stages of life in rat. Neurosci. Biobehav. Rev. 2012, 36, 1463–1480. [Google Scholar] [CrossRef] [PubMed]
- Repo-Tiihonen, E.; Halonen, P.; Tiihonen, J.; Virkkunen, M. Total serum cholesterol level, violent criminal offences, suicidal behavior, mortality and the appearance of conduct disorder in finnish male criminal offenders with antisocial personality disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2002, 252, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Virkkunen, M.; Rissanen, A.; Franssila-Kallunki, A.; Tiihonen, J. Low non-oxidative glucose metabolism and violent offending: An 8-year prospective follow-up study. Psychiatry Res. 2009, 168, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Bjork, J.M.; Dougherty, D.M.; Moeller, F.G.; Cherek, D.R.; Swann, A.C. The effects of tryptophan depletion and loading on laboratory aggression in men: Time course and a food-restricted control. Psychopharmacology (Berl.) 1999, 142, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Virkkunen, M.; Goldman, D.; Nielsen, D.A.; Linnoila, M. Low brain serotonin turnover rate (low CSF 5-HIAA) and impulsive violence. J. Psychiatry Neurosci. 1995, 20, 271–275. [Google Scholar] [PubMed]
- Werbach, M. Nutritional influences on aggressive behavior. J. Orthomol. Med. 1992, 7, 45–51. [Google Scholar]
- Krakowski, M. Violence and serotonin: Influence of impulse control, affect regulation, and social functioning. J. Neuropsychiatry Clin. Neurosci. 2003, 15, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, D. Biobehavioral perspective in criminology. In The Wadsworth Series in Criminological Theory; Wadsworth/Thomson Learing: Belmont, CA, USA, 2001. [Google Scholar]
- Tu, J.B.; Shafey, H.; VanDewetering, C. Iron deficiency in two adolescents with conduct, dysthymic and movement disorders. Can. J. Psychiatr. Revue Can. Psychiatry 1994, 39, 371–375. [Google Scholar]
- Sever, Y.; Ashkenazi, A.; Tyano, S.; Weizman, A. Iron treatment in children with attention deficit hyperactivity disorder. A preliminary report. Neuropsychobiology 1997, 35, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Konofal, E.; Cortese, S.; Lecendreux, M.; Arnulf, I.; Mouren, M.C. Effectiveness of iron supplementation in a young child with attention-deficit/hyperactivity disorder. Pediatrics 2005, 116, e732–e734. [Google Scholar] [CrossRef] [PubMed]
- Rosen, G.M.; Deinard, A.S.; Schwartz, S.; Smith, C.; Stephenson, B.; Grabenstein, B. Iron deficiency among incarcerated juvenile delinquents. J. Adolesc. Health Care 1985, 6, 419–423. [Google Scholar] [CrossRef]
- Corapci, F.; Calatroni, A.; Kaciroti, N.; Jimenez, E.; Lozoff, B. Longitudinal evaluation of externalizing and internalizing behavior problems following iron deficiency in infancy. J. Pediatr. Psychol. 2010, 35, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Yorbik, O.; Ozdag, M.F.; Olgun, A.; Senol, M.G.; Bek, S.; Akman, S. Potential effects of zinc on information processing in boys with attention deficit hyperactivity disorder. Prog. Neuropsychopharm. Biol. Psychiatry 2008, 32, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.E.; Bozzolo, H.; Hollway, J.; Cook, A.; DiSilvestro, R.A.; Bozzolo, D.R.; Crowl, L.; Ramadan, Y.; Williams, C. Serum zinc correlates with parent- and teacher-rated inattention in children with attention-deficit/hyperactivity disorder. J. Child. Adolesc. Psychopharmacol. 2005, 15, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Steenweg-de Graaff, J.; Roza, S.J.; Steegers, E.A.; Hofman, A.; Verhulst, F.C.; Jaddoe, V.W.; Tiemeier, H. Maternal folate status in early pregnancy and child emotional and behavioral problems: The generation R study. Am. J. Clin. Nutr. 2012, 95, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Suren, P.; Roth, C.; Bresnahan, M.; Haugen, M.; Hornig, M.; Hirtz, D.; Lie, K.K.; Lipkin, W.I.; Magnus, P.; Reichborn-Kjennerud, T.; et al. Association between maternal use of folic acid supplements and risk of autism spectrum disorders in children. JAMA 2013, 309, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Muntjewerff, J.W.; van der Put, N.; Eskes, T.; Ellenbroek, B.; Steegers, E.; Blom, H.; Zitman, F. Homocysteine metabolism and B-vitamins in schizophrenic patients: Low plasma folate as a possible independent risk factor for schizophrenia. Psychiatry Res. 2003, 121, 1–9. [Google Scholar] [CrossRef]
- Hibbeln, J.R.; Ferguson, T.A.; Blasbalg, T.L. Omega-3 fatty acid deficiencies in neurodevelopment, aggression and autonomic dysregulation: Opportunities for intervention. Int. Rev. Psychiatry 2006, 18, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Buydens-Branchey, L.; Branchey, M.; Hibbeln, J.R. Associations between increases in plasma n-3 polyunsaturated fatty acids following supplementation and decreases in anger and anxiety in substance abusers. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Raine, A.; Portnoy, J.; Liu, J.; Mahoomed, T.; Hibbeln, J.R. Reduction in behavior problems with omega-3 supplementation in children aged 8–16 years: A randomized, double-blind, placebo-controlled, stratified, parallel-group trial. J. Child. Psychol. Psychiatry 2015, 56, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Walsh, W.J.; Isaacson, H.R.; Rehman, F.; Hall, A. Elevated blood copper/zinc ratios in assaultive young males. Physiol. Behav. 1997, 62, 327–329. [Google Scholar] [CrossRef]
- Hubbs-Tait, L.; Nation, J.R.; Krebs, N.F.; Bellinger, D.C. Neurotoxicants, micronutrients, and social environments individual and combined effects on childrenʼs development. Psychol. Sci. Public Interest 2005, 6, 57–121. [Google Scholar] [CrossRef] [PubMed]
- Bogden, J.D.; Oleske, J.M.; Louria, D.B. Lead poisoning—One approach to a problem that won’t go away. Environ. Health Perspect. 1997, 105, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- Woolf, A.D.; Goldman, R.; Bellinger, D.C. Update on the clinical management of childhood lead poisoning. Pediatr. Clin. N. Am. 2007, 54, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Masters, R.D.; Hone, B.; Doshi, A. Environmental pollution, neurotoxicity, and criminal violence. Environ. Toxicol. 1998, 7, 13–48. [Google Scholar]
- Needleman, H.L.; Riess, J.A.; Tobin, M.J.; Biesecker, G.E.; Greenhouse, J.B. Bone lead levels and delinquent behavior. JAMA 1996, 275, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Wang, W.; McCauley, L.; Pinto-Martin, J.; Wang, Y.; Li, L.; Yan, C.; Rogan, W.J. Blood lead concentrations and childrenʼs behavioral and emotional problems: A cohort study. JAMA Pediatr. 2014, 168, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Wolffe, A.P.; Matzke, M.A. Epigenetics: Regulation through repression. Science 1999, 286, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Heard, E.; Martienssen, R.A. Transgenerational epigenetic inheritance: Myths and mechanisms. Cell 2014, 157, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Deans, C.; Maggert, K.A. What do you mean, “epigenetic”? Genetics 2015, 199, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Radford, E.J.; Ito, M.; Shi, H.; Corish, J.A.; Yamazawa, K.; Isganaitis, E.; Seisenberger, S.; Hore, T.A.; Reik, W.; Erkek, S.; et al. In utero effects. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 2014, 345. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O.; Meaney, M.J.; Szyf, M. Diet and the epigenetic (re)programming of phenotypic differences in behavior. Brain Res. 2008, 1237, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.; Rajan, S.; Paul, J.; Andrews, S. Efficacy and safety of intrathecal pentazocine as a sole anesthetic agent for lower limb surgeries. Anesth. Essays Res. 2013, 7, 49–53. [Google Scholar] [PubMed]
- Pogribny, I.P.; Karpf, A.R.; James, S.R.; Melnyk, S.; Han, T.; Tryndyak, V.P. Epigenetic alterations in the brains of fisher 344 rats induced by long-term administration of folate/methyl-deficient diet. Brain Res. 2008, 1237, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, S.W.; Brons, C.; Bluck, L.; Hjort, L.; Faerch, K.; Thankamony, A.; Gillberg, L.; Friedrichsen, M.; Dunger, D.B.; Vaag, A.A. Metabolic response to 36 hours of fasting in young men born small vs appropriate for gestational age. Diabetologia 2015, 58, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Kundaje, A.; Meuleman, W.; Ernst, J.; Bilenky, M.; Yen, A.; Heravi-Moussavi, A.; Kheradpour, P.; Zhang, Z.; Wang, J.; Ziller, M.J.; et al. Integrative analysis of 111 reference human epigenomes. Nature 2015, 518, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Santi, D.V. On the mechanism and inhibition of DNA cytosine methyltransferases. Prog. Clin. Biol. Res. 1985, 198, 119–129. [Google Scholar] [PubMed]
- Niculescu, M.D.; Zeisel, S.H. Diet, methyl donors and DNA methylation: Interactions between dietary folate, methionine and choline. J. Nutr. 2002, 132, 2333s–2335s. [Google Scholar] [PubMed]
- Dominguez-Salas, P.; Moore, S.E.; Baker, M.S.; Bergen, A.W.; Cox, S.E.; Dyer, R.A.; Fulford, A.J.; Guan, Y.; Laritsky, E.; Silver, M.J.; et al. Maternal nutrition at conception modulates DNA methylation of human metastable epialleles. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Grissom, N.M.; Reyes, T.M. Gestational overgrowth and undergrowth affect neurodevelopment: Similarities and differences from behavior to epigenetics. Int. J. Dev. Neurosci. 2013, 31, 406–414. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Lupu, D.S.; Niculescu, M.D. Perinatal α-linolenic acid availability alters the expression of genes related to memory and to epigenetic machinery, and the Mecp2 DNA methylation in the whole brain of mouse offspring. Int. J. Dev. Neurosci. 2014, 36, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Langie, S.A.; Achterfeldt, S.; Gorniak, J.P.; Halley-Hogg, K.J.; Oxley, D.; van Schooten, F.J.; Godschalk, R.W.; McKay, J.A.; Mathers, J.C. Maternal folate depletion and high-fat feeding from weaning affects DNA methylation and DNA repair in brain of adult offspring. FASEB J. 2013, 27, 3323–3334. [Google Scholar] [CrossRef] [PubMed]
- Ross, L.J.; Wilson, M.; Banks, M.; Rezannah, F.; Daglish, M. Prevalence of malnutrition and nutritional risk factors in patients undergoing alcohol and drug treatment. Nutrition 2012, 28, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Marjonen, H.; Sierra, A.; Nyman, A.; Rogojin, V.; Grohn, O.; Linden, A.M.; Hautaniemi, S.; Kaminen-Ahola, N. Early maternal alcohol consumption alters hippocampal DNA methylation, gene expression and volume in a mouse model. PLoS ONE 2015, 10, e0124931. [Google Scholar] [CrossRef] [PubMed]
- Eells, J.T.; Gonzalez-Quevedo, A.; Santiesteban Freixas, R.; McMartin, K.E.; Sadun, A.A. Folic acid deficiency and increased concentrations of formate in serum and cerebrospinal fluid of patients with epidemic optical neuropathy. Rev. Cubana Med. Trop. 2000, 52, 21–23. [Google Scholar] [PubMed]
- Susser, E.S.; Lin, S.P. Schizophrenia after prenatal exposure to the dutch hunger winter of 1944–1945. Arch. Gen. Psychiatry 1992, 49, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Susser, E.; Neugebauer, R.; Hoek, H.W.; Brown, A.S.; Lin, S.; Labovitz, D.; Gorman, J.M. Schizophrenia after prenatal famine. Further evidence. Arch. Gen. Psychiatry 1996, 53, 25–31. [Google Scholar] [CrossRef] [PubMed]
- St Clair, D.; Xu, M.; Wang, P.; Yu, Y.; Fang, Y.; Zhang, F.; Zheng, X.; Gu, N.; Feng, G.; Sham, P.; et al. Rates of adult schizophrenia following prenatal exposure to the Chinese famine of 1959–1961. JAMA 2005, 294, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Slagboom, P.E.; van Dongen, J.; Kremer, D.; Stein, A.D.; Putter, H.; Heijmans, B.T.; Lumey, L. Prenatal famine and genetic variation are independently and additively associated with DNA methylation at regulatory loci within IGF2/H19. PLoS ONE 2012, 7, e37933. [Google Scholar] [CrossRef] [PubMed]
- Lumey, L.H.; Terry, M.B.; Delgado-Cruzata, L.; Liao, Y.; Wang, Q.; Susser, E.; McKeague, I.; Santella, R.M. Adult global DNA methylation in relation to pre-natal nutrition. Int. J. Epidemiol. 2012, 41, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Goeman, J.J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.C.; Stok, A.P.; Thijssen, P.E.; Muller, F.; et al. DNA methylation signatures link prenatal famine exposure to growth and metabolism. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Tsankova, N.; Renthal, W.; Kumar, A.; Nestler, E.J. Epigenetic regulation in psychiatric disorders. Nat. Rev. Neurosci. 2007, 8, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Cervoni, N.; Detich, N.; Seo, S.B.; Chakravarti, D.; Szyf, M. The oncoprotein Set/TAF-1β, an inhibitor of histone acetyltransferase, inhibits active demethylation of DNA, integrating DNA methylation and transcriptional silencing. J. Biol. Chem. 2002, 277, 25026–25031. [Google Scholar] [CrossRef] [PubMed]
- Vucetic, Z.; Totoki, K.; Schoch, H.; Whitaker, K.W.; Hill-Smith, T.; Lucki, I.; Reyes, T.M. Early life protein restriction alters dopamine circuitry. Neuroscience 2010, 168, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.; Goyal, D.; Leitzke, A.; Gheorghe, C.P.; Longo, L.D. Brain renin-angiotensin system: Fetal epigenetic programming by maternal protein restriction during pregnancy. Reprod. Sci. 2010, 17, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Radlowski, E.C.; Johnson, R.W. Perinatal iron deficiency and neurocognitive development. Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.V.; Kennedy, B.C.; Lien, Y.C.; Simmons, R.A.; Georgieff, M.K. Fetal iron deficiency induces chromatin remodeling at the BDNF locus in adult rat hippocampus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R276–R282. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sharma, R.P.; Costa, R.H.; Costa, E.; Grayson, D.R. On the epigenetic regulation of the human reelin promoter. Nucleic Acids Res. 2002, 30, 2930–2939. [Google Scholar] [CrossRef] [PubMed]
- Niculescu, M.D.; Yamamuro, Y.; Zeisel, S.H. Choline availability modulates human neuroblastoma cell proliferation and alters the methylation of the promoter region of the cyclin-dependent kinase inhibitor 3 gene. J. Neurochem. 2004, 89, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Grissom, N.M.; Herdt, C.T.; Desilets, J.; Lidsky-Everson, J.; Reyes, T.M. Dissociable deficits of executive function caused by gestational adversity are linked to specific transcriptional changes in the prefrontal cortex. Neuropsychopharmacology 2015, 40, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Onishchenko, N.; Karpova, N.; Sabri, F.; Castren, E.; Ceccatelli, S. Long-lasting depression-like behavior and epigenetic changes of BDNF gene expression induced by perinatal exposure to methylmercury. J. Neurochem. 2008, 106, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Nakagawasai, O.; Yamadera, F.; Sato, S.; Taniguchi, R.; Hiraga, H.; Arai, Y.; Murakami, H.; Mawatari, K.; Niijima, F.; Tan-No, K.; et al. Alterations in cognitive function in prepubertal mice with protein malnutrition: Relationship to changes in choline acetyltransferase. Behav. Brain Res. 2006, 167, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Sara, V.R.; King, T.L.; Lazarus, L. The influence of early nutrition and environmental rearing on brain growth and behaviour. Experientia 1976, 32, 1538–1540. [Google Scholar] [CrossRef] [PubMed]
- Gressens, P.; Muaku, S.M.; Besse, L.; Nsegbe, E.; Gallego, J.; Delpech, B.; Gaultier, C.; Evrard, P.; Ketelslegers, J.M.; Maiter, D. Maternal protein restriction early in rat pregnancy alters brain development in the progeny. Brain Res. Dev. Brain Res. 1997, 103, 21–35. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Naninck, E.F.; van Goudoever, J.B.; Fitzsimons, C.; Joels, M.; Korosi, A. Perinatal programming of adult hippocampal structure and function; emerging roles of stress, nutrition and epigenetics. Trends Neurosci. 2013, 36, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Shoham, S.; Youdim, M.B. The effects of iron deficiency and iron and zinc supplementation on rat hippocampus ferritin. J. Neural Transm. 2002, 109, 1241–1256. [Google Scholar] [CrossRef] [PubMed]
- Black, M.M.; Baqui, A.H.; Zaman, K.; Ake Persson, L.; El Arifeen, S.; Le, K.; McNary, S.W.; Parveen, M.; Hamadani, J.D.; Black, R.E. Iron and zinc supplementation promote motor development and exploratory behavior among Bangladeshi infants. Am. J. Clin. Nutr. 2004, 80, 903–910. [Google Scholar] [PubMed]
- Colombo, J.; Kannass, K.N.; Shaddy, D.J.; Kundurthi, S.; Maikranz, J.M.; Anderson, C.J.; Blaga, O.M.; Carlson, S.E. Maternal DHA and the development of attention in infancy and toddlerhood. Child. Dev. 2004, 75, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Glenn, M.J.; Kirby, E.D.; Gibson, E.M.; Wong-Goodrich, S.J.; Mellott, T.J.; Blusztajn, J.K.; Williams, C.L. Age-related declines in exploratory behavior and markers of hippocampal plasticity are attenuated by prenatal choline supplementation in rats. Brain Res. 2008, 1237, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Novak, E.M.; Dyer, R.A.; Innis, S.M. High dietary omega-6 fatty acids contribute to reduced docosahexaenoic acid in the developing brain and inhibit secondary neurite growth. Brain Res. 2008, 1237, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Raine, A.; Venables, P.H.; Dalais, C.; Mednick, S.A. Malnutrition at age 3 years and lower cognitive ability at age 11 years: Independence from psychosocial adversity. Arch. Pediatr. Adolesc. Med. 2003, 157, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Wutman, R.J. Ways That Foods Can Affect the Brain; Guilford Press: New York, NY, USA, 1990. [Google Scholar]
- Raine, A.; Yang, Y.; Narr, K.L.; Toga, A.W. Sex differences in orbitofrontal gray as a partial explanation for sex differences in antisocial personality. Mol. Psychiatry 2011, 16, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yasuda, A.; Kawazato, H.; Sakai, K.; Shimada, T.; Takeshita, M.; Yuasa, S.; Kobayashi, T.; Watanabe, S.; Okuyama, H. Synaptic vesicle ultrastructural changes in the rat hippocampus induced by a combination of α-linolenate deficiency and a learning task. J. Neurochem. 1997, 68, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Kodas, E.; Galineau, L.; Bodard, S.; Vancassel, S.; Guilloteau, D.; Besnard, J.C.; Chalon, S. Serotoninergic neurotransmission is affected by n-3 polyunsaturated fatty acids in the rat. J. Neurochem. 2004, 89, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Turiak, G.; Galler, J.; Volicer, L. Postnatal changes of brain monoamine levels in prenatally malnourished and control rats. Int. J. Dev. Neurosci. 1997, 15, 257–263. [Google Scholar] [CrossRef]
- Marichich, E.S.; Molina, V.A.; Orsingher, O.A. Persistent changes in central catecholaminergic system after recovery of perinatally undernourished rats. J. Nutr. 1979, 109, 1045–1050. [Google Scholar] [PubMed]
- Palmer, A.A.; Brown, A.S.; Keegan, D.; Siska, L.D.; Susser, E.; Rotrosen, J.; Butler, P.D. Prenatal protein deprivation alters dopamine-mediated behaviors and dopaminergic and glutamatergic receptor binding. Brain Res. 2008, 1237, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Belluscio, L.M.; Berardino, B.G.; Ferroni, N.M.; Ceruti, J.M.; Canepa, E.T. Early protein malnutrition negatively impacts physical growth and neurological reflexes and evokes anxiety and depressive-like behaviors. Physiol. Behav. 2014, 129, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Halperin, J.M.; Sharma, V.; Siever, L.J.; Schwartz, S.T.; Matier, K.; Wornell, G.; Newcorn, J.H. Serotonergic function in aggressive and nonaggressive boys with attention deficit hyperactivity disorder. Am. J. Psychiatry 1994, 151, 243–248. [Google Scholar] [PubMed]
- Kyes, R.C.; Botchin, M.B.; Kaplan, J.R.; Manuck, S.B.; Mann, J.J. Aggression and brain serotonergic responsivity: Response to slides in male macaques. Physiol. Behav. 1995, 57, 205–208. [Google Scholar] [CrossRef]
- Weiser, M.; Levkowitch, Y.; Neuman, M.; Yehuda, S. Decrease of serum iron in acutely psychotic schizophrenic patients. Int. J. Neurosci. 1994, 78, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, L.; Delpal, S.; Guilloteau, D.; Aioun, J.; Durand, G.; Chalon, S. Chronic n-3 polyunsaturated fatty acid deficiency alters dopamine vesicle density in the rat frontal cortex. Neurosci. Lett. 2000, 284, 25–28. [Google Scholar] [CrossRef]
- Arnold, L.E.; Pinkham, S.M.; Votolato, N. Does zinc moderate essential fatty acid and amphetamine treatment of attention-deficit/hyperactivity disorder? J. Child. Adolesc. Psychopharmacol. 2000, 10, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.C.; Chen, K. MAO-A and -B gene knock-out mice exhibit distinctly different behavior. Neurobiology 1999, 7, 235–246. [Google Scholar] [PubMed]
- Schalling, D.; Asberg, M.; Edman, G.; Oreland, L. Markers for vulnerability to psychopathology: Temperament traits associated with platelet MAO activity. Acta Psychiatr. Scand. 1987, 76, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Stalenheim, E.G.; von Knorring, L.; Oreland, L. Platelet monoamine oxidase activity as a biological marker in a Swedish forensic psychiatric population. Psychiatry Res. 1997, 69, 79–87. [Google Scholar] [CrossRef]
- Caspi, A.; McClay, J.; Moffitt, T.E.; Mill, J.; Martin, J.; Craig, I.W.; Taylor, A.; Poulton, R. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Shaik, A.P.; Jamil, K. A study on the ALAD gene polymorphisms associated with lead exposure. Toxicol. Ind. Health 2008, 24, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, M.; Wang, S.; Yang, X.; Zhou, S.; Zhang, J.; Liu, Q.; Sun, Y. Lead exposure suppressed ALAD transcription by increasing methylation level of the promoter cpg islands. Toxicol. Lett. 2011, 203, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Tasmin, S.; Furusawa, H.; Ahmad, S.A.; Faruquee, M.H.; Watanabe, C. Delta-aminolevulinic acid dehydratase (ALAD) polymorphism in lead exposed bangladeshi children and its effect on urinary aminolevulinic acid (ALA). Environ. Res. 2015, 136, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Cecil, K.M.; Brubaker, C.J.; Adler, C.M.; Dietrich, K.N.; Altaye, M.; Egelhoff, J.C.; Wessel, S.; Elangovan, I.; Hornung, R.; Jarvis, K.; et al. Decreased brain volume in adults with childhood lead exposure. PLoS Med. 2008, 5, e112. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Anderson, D.W.; Wade, T.V.; Smith, M.G.; Leibrandt, P.; Zuck, L.; Lidsky, T.I. Inhibition of progenitor cell proliferation in the dentate gyrus of rats following post-weaning lead exposure. Neurotoxicology 2005, 26, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Ponzoni, S.; Guimaraes, F.S.; del Bel, E.A.; Garcia-Cairasco, N. Behavioral effects of intra-nigral microinjections of manganese chloride: Interaction with nitric oxide. Prog. Neuropsychopharmacol. Biol. Psychiatry 2000, 24, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Raine, A.; Buchsbaum, M.; LaCasse, L. Brain abnormalities in murderers indicated by positron emission tomography. Biol. Psychiatry 1997, 42, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Delville, Y. Exposure to lead during development alters aggressive behavior in golden hamsters. Neurotoxicol. Teratol. 1999, 21, 445–449. [Google Scholar] [CrossRef]
- Burns, J.M.; Baghurst, P.A.; Sawyer, M.G.; McMichael, A.J.; Tong, S.L. Lifetime low-level exposure to environmental lead and childrenʼs emotional and behavioral development at ages 11–13 years. The port pirie cohort study. Am. J. Epidemiol. 1999, 149, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.N.; Ris, M.D.; Succop, P.A.; Berger, O.G.; Bornschein, R.L. Early exposure to lead and juvenile delinquency. Neurotoxicol. Teratol. 2001, 23, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.P.; Dietrich, K.N.; Ris, M.D.; Hornung, R.W.; Wessel, S.D.; Lanphear, B.P.; Ho, M.; Rae, M.N. Association of prenatal and childhood blood lead concentrations with criminal arrests in early adulthood. PLoS Med. 2008, 5, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Murphy, V.A.; Rosenberg, J.M.; Smith, Q.R.; Rapoport, S.I. Elevation of brain manganese in calcium-deficient rats. Neurotoxicology 1991, 12, 255–263. [Google Scholar] [PubMed]
- Nihei, M.K.; Desmond, N.L.; McGlothan, J.L.; Kuhlmann, A.C.; Guilarte, T.R. N-methyl-d-aspartate receptor subunit changes are associated with lead-induced deficits of long-term potentiation and spatial learning. Neuroscience 2000, 99, 233–242. [Google Scholar] [CrossRef]
- Szyf, M. Towards a pharmacology of DNA methylation. Trends Pharmacol. Sci. 2001, 22, 350–354. [Google Scholar] [CrossRef]
- Bolton, J.L.; Bilbo, S.D. Developmental programming of brain and behavior by perinatal diet: Focus on inflammatory mechanisms. Dialogues Clin. Neurosci. 2014, 16, 307–320. [Google Scholar] [PubMed]
- Paquette, A.G.; Lesseur, C.; Armstrong, D.A.; Koestler, D.C.; Appleton, A.A.; Lester, B.M.; Marsit, C.J. Placental HTR2A methylation is associated with infant neurobehavioral outcomes. Epigenetics 2013, 8, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Pan, Y.X. Pathophysiological basis for compromised health beyond generations: Role of maternal high-fat diet and low-grade chronic inflammation. J. Nutr. Biochem. 2015, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Raine, A. Biosocial studies of antisocial and violent behavior in children and adults: A review. J. Abnorm. Child. Psychol. 2002, 30, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, R.E.; Nagin, D.S.; Seguin, J.R.; Zoccolillo, M.; Zelazo, P.D.; Boivin, M.; Perusse, D.; Japel, C. Physical aggression during early childhood: Trajectories and predictors. Pediatrics 2004, 114, e43–e50. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhao, S.R.; Reyes, T. Neurological and Epigenetic Implications of Nutritional Deficiencies on Psychopathology: Conceptualization and Review of Evidence. Int. J. Mol. Sci. 2015, 16, 18129-18148. https://doi.org/10.3390/ijms160818129
Liu J, Zhao SR, Reyes T. Neurological and Epigenetic Implications of Nutritional Deficiencies on Psychopathology: Conceptualization and Review of Evidence. International Journal of Molecular Sciences. 2015; 16(8):18129-18148. https://doi.org/10.3390/ijms160818129
Chicago/Turabian StyleLiu, Jianghong, Sophie R. Zhao, and Teresa Reyes. 2015. "Neurological and Epigenetic Implications of Nutritional Deficiencies on Psychopathology: Conceptualization and Review of Evidence" International Journal of Molecular Sciences 16, no. 8: 18129-18148. https://doi.org/10.3390/ijms160818129