Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest


1
Centre for Integrative Ecology, School of Life and Environmental Sciences, Deakin University, Burwood, Victoria 3125, Australia
2
Department of Forestry, College of Forest Resources, Mississippi State University, Mississippi State, MS 39762, USA
*
Author to whom correspondence should be addressed.
Forests 2019, 10(12), 1121; https://doi.org/10.3390/f10121121
Submission received: 28 October 2019
/
Revised: 4 December 2019
/
Accepted: 5 December 2019
/
Published: 9 December 2019
(This article belongs to the Special Issue Forests Carbon and Water Dynamics)
Abstract
:Oaks (Quercus) are a dominant forest species throughout much of the eastern United States. However, oak regeneration failure due to a myriad of issues (e.g., suppression of natural fire, excess nitrogen deposition, pressure from herbivore activity) is leading to a decline in oak dominance. This change may alter forest hydrology and nutrients through variation in species characteristics. Throughfall (TF) and stemflow (SF) quantity and chemistry were sampled during storm events under oak and non-oak (hickory, Carya) species to quantify differences in canopy-derived water and nutrients from an upland oak-hickory forest in Mississippi. Stemflow partitioning was 86% higher in hickory species compared to oak species (394.50 L m−2; p < 0.001). Across all species, dissolved organic carbon (DOC) was 1.5 times greater in throughfall (p = 0.024) and 8.7 times greater in stemflow (p < 0.001) compared to rainfall. White oak DOC concentrations (TF: 22.8 ± 5.5 mg L−1; SF: 75.1 ± 9.5 mg L−1) were greater compared to hickory species (TF: 21.0 ± 18.3 mg L−1; SF: 34.5 ± 21.0 mg L−1) (p = 0.048). Results show that while smoother-barked hickory species generate more stemflow volume, rougher-barked oak species generate stemflow that is more enriched in nutrients, which is a function of the canopy characteristics of each species. Within a single stand, this study demonstrates how variable water and nutrient fluxes may be and provide insights into species-level variability in oak-hickory forest types that may be undergoing compositional changes.
1. Introduction
Oaks (Quercus) are a dominant overstory species in many upland forest ecosystems across the eastern United States that contribute vital ecosystem services through water and nutrient cycling [1,2,3,4,5]. Given the prevalence, longevity, and diversity of oak species in forest ecosystems, it is likely that these species strongly mediate nutrient cycling when present [6,7,8]. During the past century, the decline of upland oak species due to a myriad of issues (e.g., suppression of natural fire [7,9], excess nitrogen deposition [10,11], pressure from herbivore activity [12,13], climate change [14]) has led to a shift in species composition in temperate deciduous forests from shade-intolerant, xeric species (i.e., oak) to more shade-tolerant, mesic species (i.e., ‘mesophytes’). Red maple (Acer rubrum L.) has garnered the most attention as a mesophyte across the Central Hardwoods region since the term’s coinage in 2008 [15]. However, recent work has considered other shade-tolerant/fire-sensitive species, including sweetgum (Liquidambar styraciflua L.), winged elm (Ulmus alata Michx.), Florida maple (Acer floridanum (Chapm.) Pax), American beech (Fagus grandifolia Ehrh.), and upland hickories (Carya spp.) [16,17,18,19]. In the southeastern portion of the United States, increased understory growth due to fire suppression has created cooler and more humid understory microclimates [15]. This inhibits the regeneration of upland oaks, which require disturbance (i.e., fire) to increase understory light availability and reduce shading from fire-intolerant and shade-tolerant species [20].
The tree canopy is the first major compartment encountered by precipitation as it moves through forested ecosystems, and can substantially transform the fate and transport of water and nutrients [21,22,23]. Precipitation that is not intercepted is partitioned by tree canopies into hydrological fluxes, such as throughfall and stemflow. Throughfall is the precipitation falling through the canopy to the forest floor and accounts for 70% to 90% of incident rainfall in temperate forests [24]. Stemflow is the precipitation captured by the canopy that is then funneled down the stem of trees and produces 3% to 10% of forest soil water inputs [25].
As a dominant canopy species, oaks are primed to contribute significant inputs of water and nutrients to the forest ecosystem through pathways, including throughfall and stemflow, that are uniquely modified by interspecific characteristics [2,6,26,27]. Throughfall and stemflow are spatially and temporally variable hydrologic fluxes that are influenced by a host of factors, including: Physiological and morphological traits related to forest composition; seasonality and the presence of foliage; precipitation characteristics; and meteorological conditions [28,29,30,31]. Nested within these hydrologic fluxes are biogeochemical fluxes that are additionally influenced by canopy characteristics, such as atmospheric scavenging capabilities and internal leaf nutrient contents [32,33,34]. Oak leaves tend to have a higher nitrogen content than other tree species [7], leading to higher rates of nitrogen leaching during rainfall events [35].
It is currently unknown how the diminishing dominance of oak species and subsequent increasing presence of shade-tolerant and mesophytic understory species will influence throughfall and stemflow hydrology and biogeochemistry in eastern deciduous forests of the United States. Therefore, the objectives of this research were to quantify and contrast the contribution of oaks versus codominant hickories to canopy-derived throughfall and stemflow hydrology and chemistry in temperate deciduous forest in Mississippi. These forests are comprised of a high diversity of species, even within the oak and hickory genus, making the selection of a few canopy-dominant species difficult. As such, this study presents hydrology and biogeochemistry data on six of the most dominant overstory species that are common throughout southeastern forests.
2. Materials and Methods
2.1. Site Description
A 15-ha experimental field site was established at Sessums Natural Area (SNA) in Starkville, Mississippi (33.42° N, 88.76° W) to monitor throughfall and stemflow hydrology and biogeochemistry over the course of a single water year from November 2014 through October 2015. Dominant canopy trees at the site include white oak (Quercus alba L.), post oak (Q. stellata Wangenh.), cherrybark oak (Q. pagoda Raf.), Shumard oak (Q. shumardii Buckland), shagbark hickory (Carya ovata (Mill.) K. Koch), and pignut hickory (C. glabra Miller). Leaf area index (LAI) of the stand was estimated to be 4.16 m2 m−2 from litter trap collections during the leaf fall periods of 2014 and 2015. The leaf area index (LAI) is the single-sided leaf area per unit ground surface area (m2 m−2). LAI was calculated by:
where FDW is the foliar dry weight (g), SLA is the specific leaf area (cm2 g−1), and z is the litter trap area (m2). Leaf litter traps (0.75 m2) were placed in SNA (10 total) to quantify the total amount of LAI. The site was located at the contact point between the Demopolis chalk formation to the northeast and the Ripley formation to the southwest. Soils at the site were silty clay loams ranging from somewhat poorly drained (Kipling) to well drained (Sumter) depending on landscape position [36]. Annual summer temperatures (JJA) range from 23.5 to 27.7 °C. Annual winter temperatures (DJF) range from 6.6 to 14.4 °C [37]. Thirty-year (1980–2010) normal precipitation is 140.3 cm that is evenly distributed throughout the year [37].
2.2. Experimental Design
Due to the high species diversity in southeastern forests, and at this site in particular, we selected a variety of oak and hickory species that constitute the majority of overstory species. Species selected for study from the Sect. Quercus, or the white oaks, were white oak and post oak. Species selected for study from the Sect. Lobatae, or the red oaks, were Shumard oak and cherrybark oak. Hickory species selected for study were pignut and shagbark hickory. Sample trees were selected based on canopy dominance within the stand. Measurements of diameter at breast height (DBH), crown area, and tree height for three trees of each study species were made in fall 2014 (Table 1). The crown area for each tree was calculated by measuring the crown’s radius at 6 different locations and taking the average of these measurements. Area calculations were made using the area formula: Canopy area = πr2, where r is the radius of the canopy. Tree height was measured using a TruPulse 360° R Laser Rangefinder (Laser Technology Inc., Centennial, CO, USA).
2.3. Stemflow, Throughfall, and Precipitation Measurements
In this study, we sampled chemistry during discrete rainfall events to isolate the difference in canopy-derived chemistry across species and across seasons. Only rainfall events that were large enough to generate stemflow were included in this investigation. During the dormant season when there was no foliage in the canopy, the threshold for stemflow generation was approximately 0.5 cm. During the growing season when foliage was present in the canopy and intercepted a large portion of rainfall, the threshold for stemflow generation was closer to 1.0 cm. Additionally, rainfall characteristics, such as intensity and duration, determine how much water the canopy may intercept, so the threshold of stemflow generation varied across sampling events. As such, field collectors were checked after each rainfall event, but only those events that produced enough stemflow from all study trees were included in this manuscript. In total, 16 rainfall events were sampled in this study.
Three dominant canopy trees of each species (Table 1) were selected to measure stemflow and throughfall. Each tree was outfitted with a stemflow collar constructed from polyethylene tubing ranging from a 2.54- to 3.81-cm inner diameter. The tubing was cut longitudinally and wrapped around the bole of the tree with aluminum nails and silicone caulk. The tubing drained into large collection containers (+100 L). Following each rainfall event, stemflow depth was measured in each container and converted to volume using container-specific regression equations that were derived in the laboratory. Water in stemflow collector bins was then stirred and a homogenous 250-mL subsample was collected for chemical analysis.
To measure throughfall, a 1-L high density polyethylene (HDPE) bottle fitted with a 20.3-cm diameter funnel was deployed under the crowns of the same trees sampled for stemflow. Each throughfall collector was placed midway between the main bole and the edge of the crown drip line to ensure that all water collected was from the designated tree crown. Placing throughfall apparatuses at the mid-canopy point ensured that throughfall collection was from the study species, thus reducing interference from surrounding tree species [26,38]. Following each rainfall event, throughfall volume was recorded directly with a graduated cylinder, stirred, then a homogenous 250-mL subsample was collected for chemical analysis. Throughfall containers were relocated beneath the crown’s midway point bimonthly to capture variation under the crowns of each species selected for observation.
Throughfall and stemflow volumes were converted to volumetric flux in liters per basal area (L/m2 of basal area). Basal area is a much more common measurement to understand the density of forest stands, particularly of individual species, and enables volume measurements to be scaled to the stand level on a per basal area basis [25]. Thus, it is much easier to derive basal area from field measurements of DBH, as opposed to canopy area, and thus presenting volumetric flux per basal area can be a more accessible metric for evaluation and management. Precipitation sampling occurred at the nearest accessible open location, approximately 3.0 km southeast of SNA at the Mississippi State University Dairy Farm. Rainfall characteristics, including amount (cm), duration (hr.), intensity (cm/hr.), and wetting/drying cycles were recorded using an RG3-M tipping bucket rain gauge (Onset Computer Corporation, Bourne, MA, USA) located at the Mississippi State University Dairy Farm. To sample precipitation chemistry, a collector consisting of a 1-L high density polyethylene bottle (HDPE) fitted with a 20.32-cm diameter funnel was deployed. Wool gauze was inserted into the neck of the funnel to prevent the entry of large particulates and collection bottles were enclosed in opaque bags to prevent algal growth, which may alter sample chemistry [39]. Precipitation samples were collected following the throughfall protocol.
2.4. Chemical Analysis
Precipitation, throughfall, and stemflow water samples were collected within 24 h of the cessation of a rainfall event, measured for pH using an AP951 Accumet pH meter (Fisher Scientific, Pittsburgh, PA, USA), and filtered to remove particulates greater than 0.45 µm. Samples were stored at 4 °C. Samples were analyzed for dissolved organic carbon (DOC) and dissolved nitrogen components (DON, NH4+, and NO3−). Inorganic nitrogen (NH4+-N and NO3−-N) was measured using wet ion chromatography (Dionex DX-500, Thermo Fisher Scientific Inc., Sunnyvale, CA, USA). DON was calculated by subtracting NH4+-N from total kjeldahl nitrogen (TKN), where TKN was obtained through persulfate digestion and colorimetry methodologies (Bran+Luebbe Autoanalyzer 3, SEAL Analytical Inc., Mequon, WI, USA). DOC was quantified by spectrometry with a HACH Low Range Total Organic Carbon Test kit and run on a HACH DR 5000 UV-Vis Spectrophotometer (Loveland, CO, USA). All samples were run in the Forest Hydrology and Soils Laboratory at Mississippi State University following quality assurance/quality check protocols outlined by the Environmental Protection Agency using standards, blanks, and duplicates.
As rainfall moves along hydrologic pathways and interacts with various surfaces of the forest, stemflow and throughfall may become enriched with solutes compared to incident rainfall. Therefore, the volumetric flux-based enrichment ratio was used to describe the ratio of throughfall or stemflow solute flux relative to the corresponding precipitation solute flux. The volumetric flux-based enrichment ratio for stemflow reveals the extent to which stemflow solute fluxes exceed the flux that would be input over the same area by open rainfall. For stemflow, enrichment ratios were calculated by:
where ESF is the enrichment ratio of stemflow, [SF] is the solute concentration in stemflow, SF is stemflow volume, [PG] is the solute concentration of incident rainfall, PG is the depth equivalent of incident rainfall, and BA is the basal area of the stemflow-generating tree [40]. For throughfall, flux-based enrichment ratios were calculated by:
where ETF is the enrichment ratio of throughfall, [TF] is the solute concentration in throughfall, TF is the depth equivalent of throughfall, [PG] is the solute concentration of incident rainfall, and PG is the depth equivalent of incident rainfall. In this study, since throughfall and rainfall were measured with the same gauge type, the area over which these hydrologic fluxes were measured were equal and a direct comparison could be made.
2.5. Statistical Analysis
Volumetric fluxes and solute concentrations in throughfall and stemflow between species and across seasons were tested for normality using Shapiro–Wilk tests and the homogeneity of variance using the Bartlett test. Due to the non-parametric nature of the data, a Kruskal–Wallis test was used to compare volumetric fluxes and solute concentrations in throughfall and stemflow between species and seasons. When statistical differences were present between species, specific differences between species pairings were determined using a post-hoc Wilcoxon signed-rank test. All analyses were performed in R [41].
3. Results
3.1. Throughfall and Stemflow Hydrology
A total of 16 rainfall events met the criteria of stemflow generation from all study trees, enabling the sampling and comparison of water and nutrients across species. The average size of rainfall events during the leafless period was 2.45 ± 1.17 cm and 1.78 ± 1.18 cm during the leafed period.
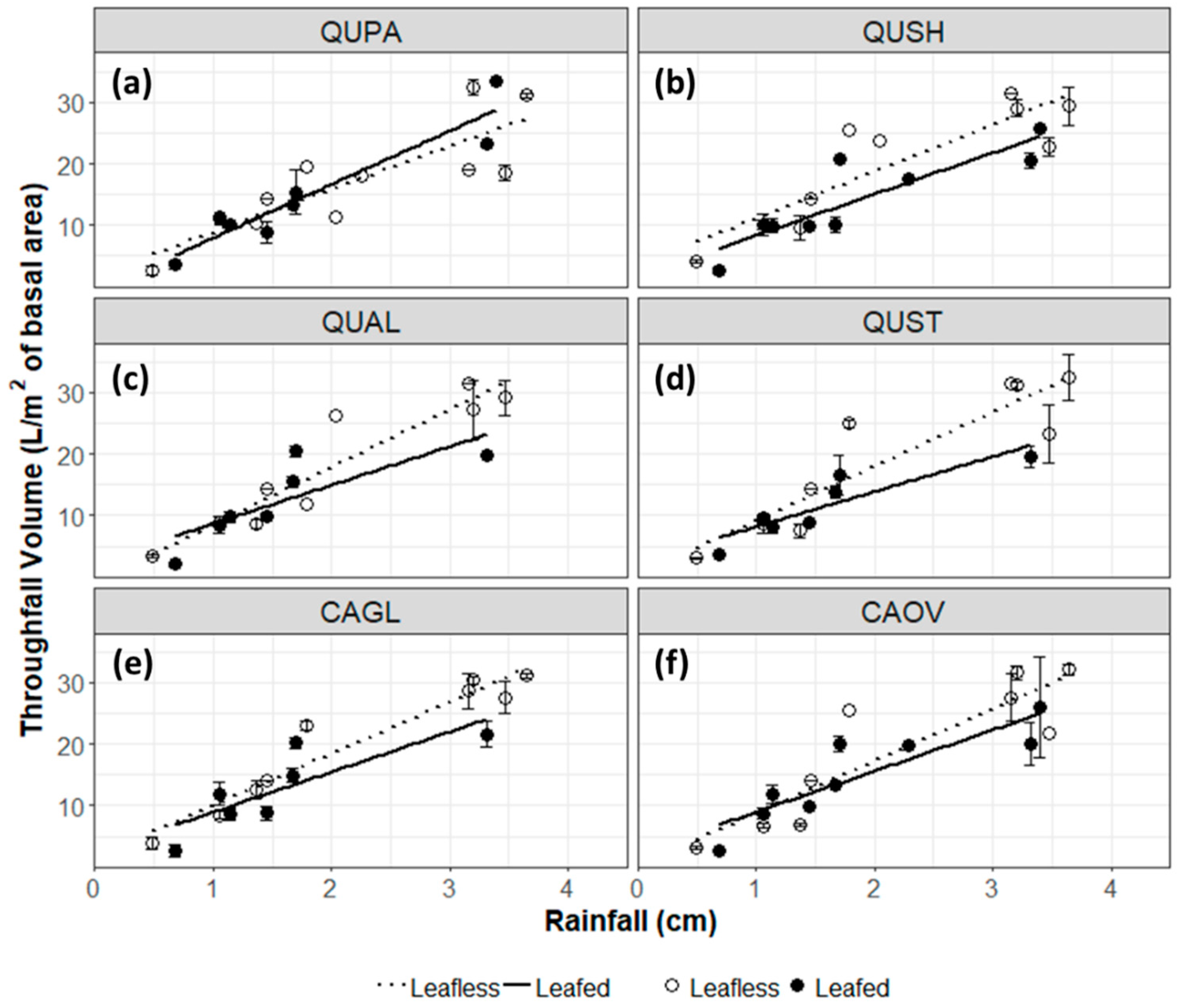
Throughfall volume per basal area across all species was largely influenced by rainfall magnitude (r2 = 0.80), with slightly stronger correlations in the leafless canopy phase (r2 = 0.800) than in the leafed (r2 = 0.76) (Figure 1). In the leafless canopy phase, the strongest correlation between rainfall and throughfall was observed under pignut hickory canopies (r2 = 0.92) and the weakest correlation under cherrybark oak canopies (r2 = 0.67). In the leafed canopy phase, the strongest correlations between rainfall and throughfall was observed in cherrybark oak (r2 = 0.87) and the weakest in white oak (r2 = 0.57). Despite these strong relationships to meteorological and environmental conditions, throughfall volumetric flux was not significantly different between species (p = 0.340) or between leaf phases (p = 0.800).
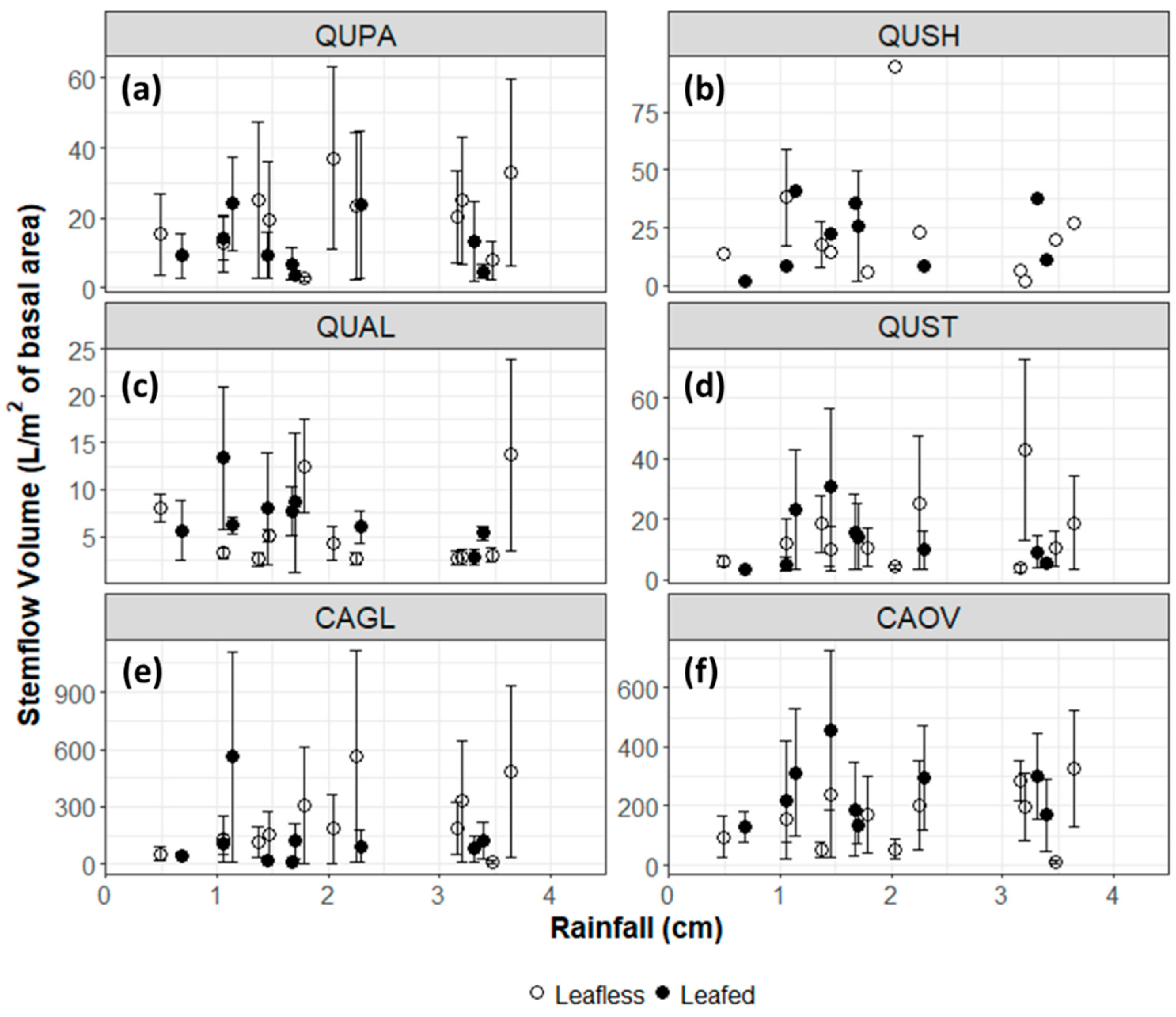
No discernable relationship was observed between stemflow volume per tree basal area as a function of rainfall amount at the annual or seasonal scale (Figure 2). However, there were significant differences in stemflow partitioning between species (p < 0.001). Per basal area, pignut hickory (CAGL) and shagbark hickory (CAOV) generated significantly more stemflow during the sampled storms than any of the oak species (p < 0.001 for all combinations, except between the two hickory species themselves). Pignut hickory had the highest average stemflow volume per basal area recorded throughout the study at 205.0 L m−2, followed by shagbark hickory (189.5 L m−2), Shumard oak (20.0 L m−2), cherrybark oak (14.8 L m−2), post oak (13.6 L m−2), and white oak (6.4 L m−2).
The combination of differences in throughfall (leafless: TF = 81.6%, leafed: TF = 74.2%, p = 0.020) and stemflow partitioning (leafless: SF = 1.2%, leafed: SF = 0.8%, p < 0.001) between seasons had counteractive effects on canopy interception, which, although larger in the leafed season, was not significantly different between seasons (leafless: INT = 17.3%, leafed: TF = 25.0%, p = 0.138) or between species (p = 0.350). The smallest increases in canopy interception from the leafless to the leafed season were observed in shagbark hickory (+4.6%, leafless = 19.1%, leafed = 23.7%) and post oak (+5.9%, leafless = 21.5%, leafed = 27.4%), with moderate increases observed in pignut hickory (+10.5%, leafless = 11.6%, leafed = 22.0%) and white oak (+11.2%, leafless = 17.1%, leafed = 28.3%), and the largest increase observed in Shumard oak (+19.2%, leafless = 13.4%, leafed = 32.5%). Surprisingly, more canopy interception was observed in cherrybark oak crowns during the leafless season than the leafed (−7.8%, leafless = 23.0%, leafed = 15.1%).
3.2. Throughfall and Stemflow DOC
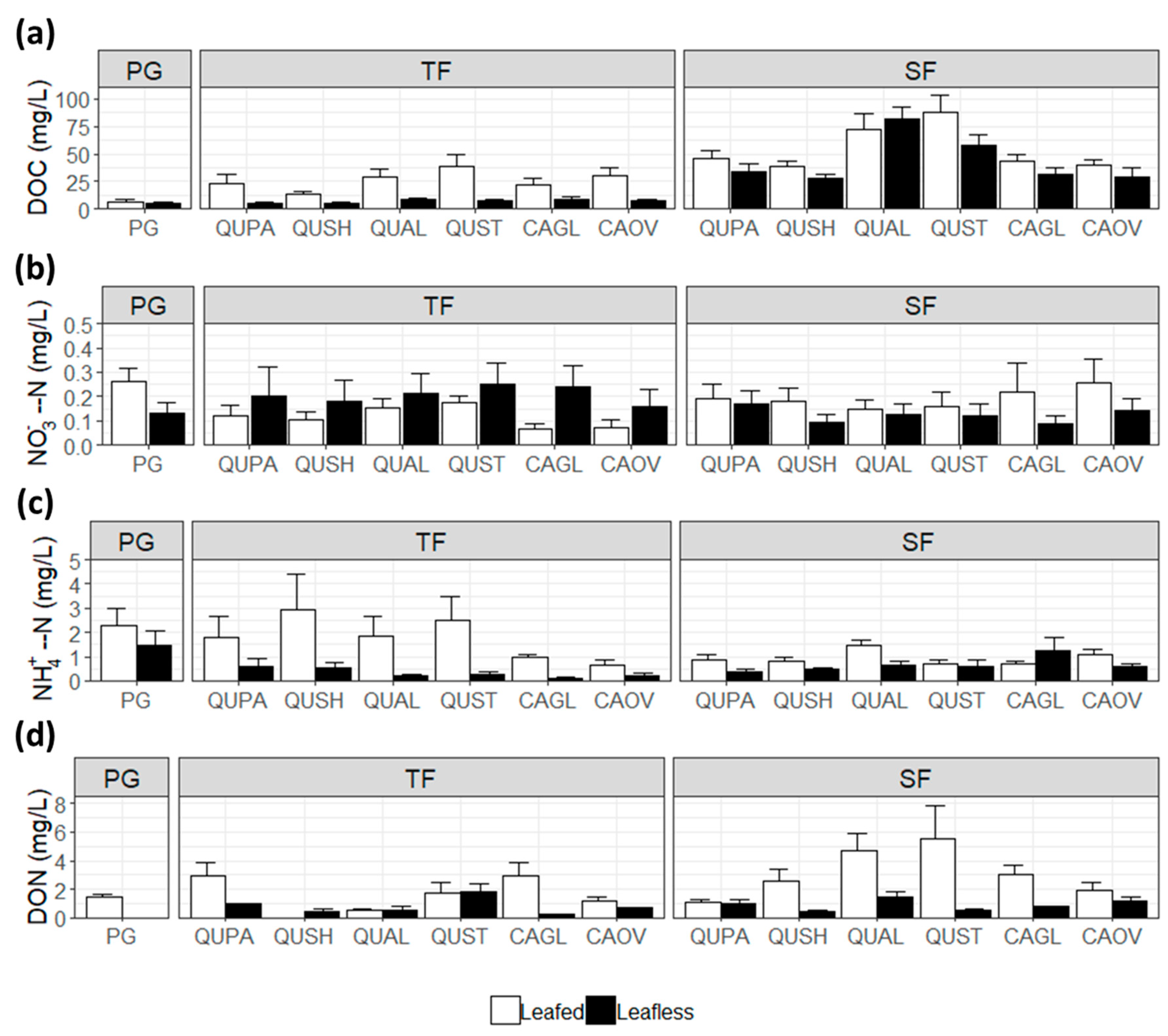
DOC concentrations were much higher beneath the canopy compared to precipitation. TF DOC was 71.6% higher than rainfall and SF DOC was 88.5% higher (TF-PG: p = 0.024 and SF-PG: p < 0.001) (Figure 3a). Further, average DOC values were 63.3% higher in TF and 31.6% higher in SF during the leafed period relative to the leafless period (p < 0.001 and p = 0.078) (Figure 3). Post oak throughfall DOC concentrations were higher compared to rainfall during the leafed season (p = 0.036), but not in the leafless season (Figure 3a). In the leafed season, pronounced differences were observed between rainfall and stemflow in white oak (p < 0.001) and post oak (p < 0.001), with borderline significant differences between rainfall and cherrybark oak (p = 0.063), as well as between shagbark hickory and post oak (p = 0.064), shagbark hickory and white oak (p = 0.080), and Shumard oak and post oak (p = 0.103) (Figure 3a). In the leafless season, white oak and post oak were again significantly different than rainfall (p < 0.001 and p = 0.014, respectively), along with differences between white oak and cherrybark oak (p = 0.020), white oak and Shumard oak (p = 0.024), white oak and pignut hickory (p = 0.013), and white oak and shagbark hickory (p = 0.048) (Figure 3a).
3.3. Throughfall and Stemflow Inorganic Nitrogen
Throughfall NO3− concentrations showed no significant differences between species (p = 0.766), precipitation (p = 0.897), and seasonality (p = 0.160). Prominent differences were observed in NO3− concentrations between all species and precipitation in stemflow (p < 0.001), although there was no seasonal variation (p = 0.604) (Figure 3b). Throughfall NH4+ concentrations showed no significant differences between species. NH4+ concentrations in throughfall were generally higher in the leafed season versus the leafless (p = 0.069). All species, with the exception of cherrybark oak (p = 0.149) and pignut hickory (p = 0.520), had significantly lower NH4+ concentrations in stemflow compared to precipitation (p < 0.050) during the leafed phase (Figure 3c). NH4+ concentrations were greater in rainfall compared to Shumard oak (p = 0.010), post oak (0.013), and shagbark hickory (p = 0.012) (Figure 3c). No distinguishable seasonal variations were detected in stemflow NH4+ concentrations (p = 0.339) (Figure 3c).
3.4. Throughfall and Stemflow Organic Nitrogen
No significant differences were observed between species, rainfall, or seasonality for DON concentrations in throughfall (TF: 4.22 ± 0.844 mg L−1; p = 0.335). Stemflow DON concentrations showed no variation between species and precipitation (SF: 3.933 ± 3.270 mg L−1; p = 0.775) (Figure 3d). Larger concentrations of DON were observed in stemflow during the leafed phase (p = 0.045), with white oak and post oak exhibiting the largest average DON concentration during the leafed season in stemflow (Figure 3d).
3.5. Throughfall and Stemflow DOC Enrichment
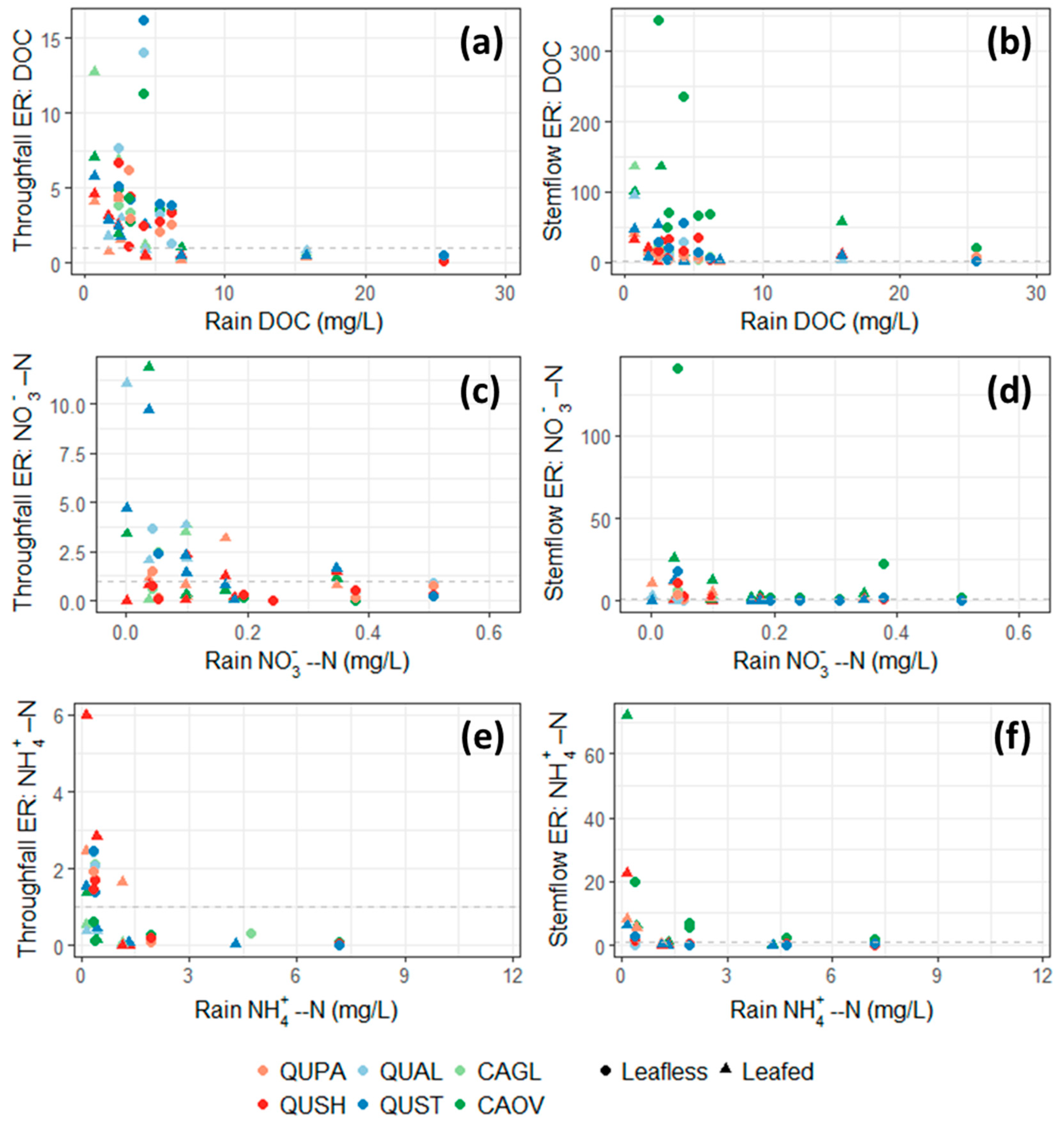
Concentration-based evaluation provides a relative method of comparing the chemical change of water in subcanopy pathways. However, to determine the overall influence, both concentration and volumetric flux must be assessed in the form of enrichment ratios. Enrichment ratios were the largest when initial solute concentrations in rainfall were small for both throughfall and stemflow (Figure 4). Average enrichment of DOC in both the leafed and leafless season was always greater than one (Table 2), signifying higher total fluxes of DOC in throughfall and stemflow relative to rainfall when differences in volumetric flux were considered. In throughfall, the highest enrichment ratios of DOC were observed under crowns of white oak and post oak (both in the white oak section) while no trend was observed among species in the leafless season (Table 2). Thus, during the growing season when foliage is present, the influence of individual species on DOC chemistry is apparent in throughfall. Additionally, throughfall DOC enrichment was generally lower in during leafless canopy conditions.
Enrichment of DOC in stemflow was an order of magnitude greater than enrichment of DOC in throughfall (Figure 4b). DOC enrichment ratios less than one were not observed in stemflow from any species. On average, stemflow DOC enrichment was slightly larger in the leafed season than the leafless, but there was no consistent trend among species (Table 2). However, in the leafless season, both hickory species had stemflow DOC enrichment ratios two to three times greater than the oak species (Table 2).
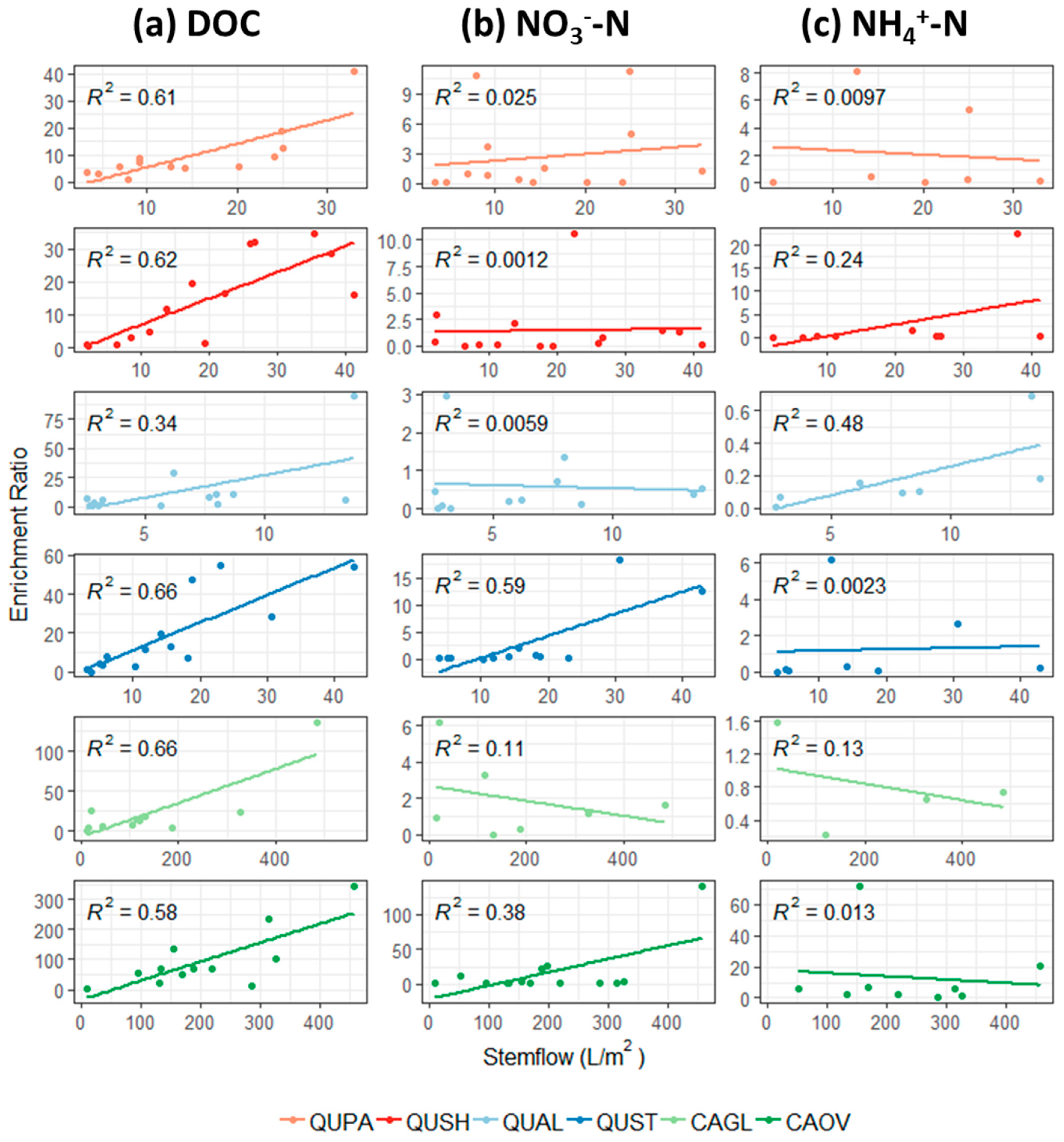
Enrichment of DOC in throughfall was positively correlated with throughfall quantity under crowns of southern red oak (R2 = 0.40) and to a smaller degree under pignut hickory (R2 = 0.27) (Figure 5a). Conversely, enrichment of DOC in stemflow was positively correlated with stemflow volume across all species (Figure 6a). The weakest correlation (R2 = 0.34) was observed between white oak stemflow volume and DOC while all other species had correlations ranging from R2 = 0.58 to 0.66.
3.6. Throughfall and Stemflow Inorganic Nitrogen Enrichment
Throughfall modifications of inorganic nitrogen were observed between seasons. For NO3−, throughfall enrichment was greater under leafless conditions compared to leafed conditions (Table 2). Conversely, NH4+ enrichment in throughfall was greater under leafed conditions compared to leafless conditions (Table 2). Approximately half of all events sampled had enrichment values for both forms of inorganic nitrogen less than one, signifying uptake/depletion of inorganic nitrogen by the forest canopy (Figure 4c,e). For NO3−, throughfall enrichment occurred predominantly under crowns of white oak and post oak (Figure 4c) while throughfall was almost always less than one under crowns of Shumard oak and cherrybark oak (Table 2). Very weak relationships were observed between throughfall volume and NO3− enrichment, with white oak, post oak, and shagbark hickory exhibiting positive correlations and pignut hickory exhibiting a negative correlation (Figure 5b). In contrast, throughfall NH4+ enrichment was negatively correlated to throughfall volume in all species (Figure 5c). Furthermore NH4+ enrichment correlations were much stronger than NO3− enrichment correlations.
Stemflow modifications of inorganic nitrogen exhibited the opposite seasonal trend as throughfall. For NO3−, stemflow enrichment was greater under leafed conditions compared to leafless conditions, and for NH4+, enrichment in stemflow was greater under leafless conditions and less than one under leafed conditions, except for Shagbark hickory (Table 2). Shagbark hickory enrichment of both forms of inorganic nitrogen was substantially larger than all other species, regardless of season (Figure 4d,f and Table 2). White oak enrichment of both forms of nitrogen was less than one regardless of season (Table 2). As stemflow volume increased, NO3− enrichment increased in post oak and shagbark hickory stemflow while NO3− enrichment decreased in pignut hickory stemflow (Figure 6b). Pignut hickory exhibit the same declining trend in NH4+ enrichment with increasing stemflow, but no trends were observed with NH4+ enrichment in post oak or shagbark hickory stemflow (Figure 6c).
4. Discussion
4.1. Throughfall and Stemflow Hydrology
Interspecific differences in throughfall and stemflow generation in this study could be attributed to varying biophysical features. The results of Marin et al. [42], for instance, found increased throughfall and stemflow hydrologic fluxes between different trees due to smaller leaf area indices and larger gap sizes. André [43] reported higher stemflow volumetric flux under beech canopies due to thinner and more angled branching compared to more horizontally developed branching in oak species. The higher stemflow partitioning beneath hickory species could be explained by similar branching patterns, where general observations in the field showed more angular and thinner branching, which could favor greater water flow on the bole of the tree. All white oaks had much rougher and thicker bark (1.53 cm) compared to all red oaks (1.03 cm) and hickories (0.91 cm), which increased bark water storage capacity [44] and may create obstacles and drip points along branches [40,45,46]. Across the spectrum of bark morphology, the six species used in this study have relatively thick and rough bark, especially compared to species, such as American beech (Fagus grandifolia Ehrh.), maple species (Acer spp.), or birch species (Betula spp.) [47]. However, in historical upland oak forests maintained by frequent, low intensity fires (or even oak-chestnut forests prior to the extirpation of Castenea dentata (Marsh.) Borkh.), the dominant, fire-tolerant species were comprised of those with thicker bark [48,49]. As such, the hickory species have relatively thinner bark than the oak species, leading to differences in water fluxes, especially with regards to stemflow. Additionally, oak species at this study site, specifically post and white oaks, hosted moss, lichen, and fern species that were present on the bole and branches of these trees (personal observation). These epiphytes not only disrupt stemflow and throughfall volumetric flux [38] but also absorb substantial quantities of water. Other studies found water absorption rates of epiphytic species to be more than 6 to 10 times their dry weight [50,51].
Stemflow is often neglected from large-scale hydrologic studies because of its minimal contribution relative to other hydrologic components. However, stemflow is an important source of water delivered directly to an individual tree’s roots. In both humid and arid regions, stemflow serves as an important and direct source of water, increasing soil moisture availability around individual root systems [23,52]. Stemflow volumetric partitioning ranged from 0.14% to 5.31% of open precipitation, which was comparable to other studies in temperate forests with stemflow measurements amounting to 10.0% of open precipitation in mixed white oak forests [53], and 7% to 8% in beech (Fagus sylvatica) forests [54,55]. Other studies noted higher stemflow conditions in young dense beech stands, which contribute more than 14% of gross precipitation [56].
The increase in average throughfall and stemflow partitioning during the leafless canopy phase is the result of the absence of vegetation during the dormant season. Lack of foliar surfaces decreases interception capacity, which limits intra-storm evaporation, thus increasing throughfall and stemflow volumetric flux. For example, Staelens et al. [57] found that throughfall volumetric fluxes decreased significantly (p = 0.014) with increasing canopy cover during the growing season. Additionally, the increased capacity for trees to generate stemflow during the leafless canopy phase has also been documented [5,31,58].
Meteorological variability has been identified as a strong control on regulating canopy-derived fluxes [30,31,59]. Storm variability has been documented to strongly influence throughfall and stemflow hydrology. Hydrologic behavior in a forest can be altered by the magnitude, duration, and intensity of precipitation [45]. Similar to studies, such as Skau [60] and Siegert et al. [61], summer rain events were shorter, higher intensity storms that were extremely isolated, and may explain the slightly higher throughfall volumetric flux during events with higher magnitude (Figure 2). In this study, general trends of hydrologic fluxes and ion concentrations were observed between some species, and it is hypothesized that more data observations would provide a clearer picture of these relationships. More replicates of the study tree species could clarify throughfall and stemflow results; however, three of each species was the largest sample size that could be accommodated given economic and practical reasons.
4.2. Throughfall and Stemflow Chemistry
4.2.1. Variations between Precipitation, Throughfall, and Stemflow
Throughfall DOC concentrations were larger in each species compared to precipitation (Figure 3), which is the result of leaching interactions from vegetative surfaces [62,63,64] and scavenging of atmospheric deposition by tree canopies. Several authors link canopy density indices, such as leaf and branch cover, to increased throughfall deposition in forests, as higher canopy density results in a greater surface area that would increase the chances of deposition capture and leaching interactions [65,66,67]. Studies show mixed results when comparing NO3− and NH4+ concentrations in forest ecosystems. André et al. [35] found significant quantities of these solutes from leafed canopies of mixed hardwoods and pure beech stands. In contrast, other studies noted a decrease of NO3− and NH4+ in throughfall due to tree, microorganism, and epiphyte uptake [68,69,70,71], which may explain the results found in this study. Further, forest canopies have been noted to capture NH4+ as dry deposition. Research suggests that NH4+ captured in forest canopies can be transformed into NO3− provided there is nitrifying bacteria present [72,73]. Other mechanisms related to canopy uptake of inorganic nitrogen species have been attributed to rainfall characteristics, including residence time of rainwater in the tree canopy [61]. These mechanisms likely occurred at the study site given the prevalence of epiphytic vegetation in the canopy (visual observation) and the variation in storms observed throughout the year-long study duration, resulting in NH4+ enrichment ratios of <1 within TF and SF. This study showed greater throughfall DOC concentrations compared to precipitation. A study examining dissolved organic matter (DOM) found similar results and determined throughfall DON and DOC concentrations derived from canopy leaching, pollen deposition, and accumulated atmospheric DOM [74]. Pollen, in particular, has the potential to strongly influence TF and SF DOC and DON concentrations in temperate deciduous forests during spring months [75]. It was noted that visibly higher pollen concentrations were observed in TF and SF samples during periods of pollen production. In fact, the highest rainfall DOC concentrations (20–30 mg/L) were observed during spring leaf emergence and pollen production in this study (Figure 4a,b).
Average annual stemflow solute concentrations were generally higher compared to precipitation, with the exception of DON. Stemflow, unlike precipitation, is provided with nutrient enrichment through foliar and stem exchange reactions as well as atmospheric deposition wash off [62,76]. Moreno et al. [77] reported higher levels of DOC in stemflow concentrations for Pyrenean oak (Quercus pyrenaica) (71.8 to 123.0 mg L−1) while concentrations in precipitation had lower DOC concentrations (5.9 to 7.1 mg L−1). Studies showed higher DON concentrations in stemflow compared to precipitation [78,79], which contrasts with the results of this study, although others have found no different between rainfall and stemflow DON [61]. The lower DON in stemflow compared to precipitation for this study may be explained by foliar or epiphytic uptake. Studies analyzing inorganic NO3− and NH4+ noted that N may be absorbed from rainfall by foliage and/or taken up by microorganisms and epiphytes [69,80]; therefore, similar patterns may describe the lower organic nitrogen content in stemflow compared to precipitation. In contrast to those studies, NO3− and NH4+ concentrations in stemflow were higher in one or more species group compared to precipitation. Inorganic N could also be leached from foliage, particularly during leaf senescence [81]. Further, the precipitation collectors could have higher nitrogen content as they were located on a dairy farm, which has the potential to expose these gauges to high nitrogen levels because of the NH3 released during farming operations [82]. Given the complex nature of the environmental nitrogen cycle, more research is required to further understand N transformations by the forest canopy.
4.2.2. Interspecific Differences in Throughfall and Stemflow Chemistry
The general trend of increased solute concentration in oak throughfall may be a result of larger canopies and canopy foliar traits [2,6,8,26,27]. In other studies, oaks had higher solute concentrations compared to other species. NH4+ and NO3− were significantly higher due to greater canopy cover in an oak-birch forest stand compared to a downy birch (Betula pubescens) swamp forest [83]. Similar to this study, Qualls and Haines [84] found that approximately 32% of the DOC entering the soil horizons originated from throughfall and stemflow, with concentrations highly dependent on vegetation type. DOC concentrations beneath white oaks were, in general, higher compared to red oak and hickory species, which may be due to the larger leaf area (Table 1), which may increase atmospheric deposition capture and leaching interactions. Variations between conifers and deciduous foliar chemistry have been linked to more DOC in soils beneath Norway spruce (Picea abies) compared to European beech (Fagus Sylitica), sessile oak (Quercus petrawa), and pedunculate oak (Quercus robur) species [85]. In our study, comparing canopy-derived chemistry solely in deciduous species, we observed general trends of greater average DON concentration in throughfall beneath white oak species compared to other species in this study. Past literature analyzing the net release of N from branches of chestnut oak (Quercus prunus) and evergreen species (Pseudotsuga spp.) yielded minimum DON leaching from the evergreen species. Phytophagous insects could also increase TF DOC and NH4+ residing in oak canopies. Oaks are generally sensitive to insect attacks, thus leached nutrients from defoliated canopies could further explain the increased TF and SF nutrient concentrations [86].
Solutes dissolved in stemflow have been documented to vary between species as well. Forest N concentrations are strongly affected by canopy interactions, as seen by this study and others [2,87]. Average annual concentrations were higher in hickory species for NH4+ and NO3− whereas white oak species had higher concentrations of both organic carbon and nitrogen. The rougher bark surface of oaks theoretically slows down stemflow and extends the residence time of the interaction between stemflow water and bark surfaces. As a result, enhanced leaching of organic forms of both nitrogen and carbon could be expected as residence time, and therefore volume, increased [88,89]. A study found lower nitrogen concentrations in stemflow to be correlated with increased epiphytic communities on various tree species in montane subtropical forests [90], suggesting uptake of inorganic nitrogen. This could explain the differences in the N concentrations between oak and hickory species. In fact, we observed a general increase in inorganic N enrichment, especially NH4+ enrichment, as stemflow increased for oak species and a general decrease in enrichment ratios of inorganic N as stemflow increased for hickory species (Figure 6). From general observations at the field site, white oak species tended to have more lichen and resurrection fern (Pleopeltis polypodioides) located on their branches. Liu et al. (2002) noted that epiphytic bryophytes decreased the annual amount of NH4+, which may determine why oak species had lower NH4+ concentrations compared to hickory species. Similar variation in DOC was observed between white birch (Betula papyrifera) and larch (Larix laricina), with reported DOC stemflow concentrations ranging from 13.74 mg L−1 in white birch to 129.91 mg L−1 in larch species [77].
4.2.3. Seasonal Differences in Throughfall and Stemflow Chemistry
The majority of solute concentrations in both throughfall and stemflow increased during the leafed phase, with the exception of NO3− and NH4+. These results correlate with findings from Lovett and Lindberg [91], where an increase of nitrogen was observed to be 25% higher during the growing season. Leaf cover increases throughfall ion concentrations by providing additional surfaces to capture atmospheric deposition that is then washed to the forest floor as throughfall during storms events [21,92,93]. Decreased concentrations of NO3− in throughfall and stemflow during the growing season were observed in this study and may be the result of the biological activity in the canopy absorbing NO3− from precipitation [62]. Additionally, physiologically active vegetation is a greater source of plant leaching than dormant vegetation [22,94,95]. In this study, significant increases in DON and DOC were observed during the leafed phase. Similar trends were observed in a hardwood forest, with solute concentrations 2 to 12 times higher during the growing season compared to the dormant season [68,78]. Other studies, however, noted greater chemical enrichment in stemflow during the winter as the reduction of foliar interception rates in deciduous forests increases the chance for wetting the bole of the tree, thus increasing the quantity of water available as stemflow to leach nutrients [62,96,97]. As such, both direct effects of seasonality (active vs. dormant canopy) and indirect effects (loss of foliage) may influence stemflow nutrient enrichment for this site.
5. Conclusions
The results of this study indicate the importance of oak foliar traits to forest hydrology, biogeochemistry, and overall forest health. It is possible then that a shift in stand composition could influence future southeastern upland oak-history forest dynamics. While our results represent one location and a single year of rainfall sampling, we provide initial evidence of hydrologic and biogeochemical differences among co-occurring species that are common throughout the southeast. Throughfall volume varied with seasonal changes in vegetation cover rather than interspecific differences between species. However, the sampling design of this study, which required sampling of only events large enough to produce stemflow (+0.5–1.0 cm), may have obscured some of the fine-scale differences in throughfall volume and chemistry that occur as smaller rainfall amounts [23]. Stemflow volume, however, was significantly different between species. Throughfall and stemflow chemistry was different than precipitation chemistry, indicating the importance of deciduous foliage in modifying precipitation chemistry. The higher solute concentrations in oak throughfall may be the result of larger crown area and canopy foliar traits, suggesting the importance that these species have to the nutrient dynamics in the upland oak-hickory forest of this study. The temporal difference between solute concentrations, with greater differences observed during the growing season among species, also supports the idea that species-level controls on forest biogeochemistry are important.
Few efforts have led to an understanding of the importance of individual species on water and nutrient cycles in high-diversity forest ecosystems undergoing compositional changes [2,23,98]. The results presented in this study have provided evidence of the importance of oak species to nutrient and water cycling in an upland oak-hickory stand in Mississippi. Broadly speaking, although oaks generate less total stemflow, the stemflow that is delivered to the forest floor is more enriched in carbon and inorganic nitrogen compared to co-occurring hickory species. As such, oaks likely play a strong role in mediating nutrient cycles in their immediate vicinity. These findings highlight the need to further understand the water and nutrient dynamics of oak-dominated ecosystems, as the current state of nutrient cycling in these forest could be altered by a shift in species dominance. Continued work that quantifies forest hydrology and biogeochemistry in oak species will add to the understanding of nutrient and water cycling for upland forests in the southeastern United States.
Author Contributions
Conceptualization, K.L. and C.S.; methodology, K.L. and C.S.; formal analysis, K.L. and C.S.; investigation, K.L.; resources, C.S.; writing—original draft preparation, K.L.; writing—review and editing, K.L. and C.S.
Funding
This work was supported by the National Institution of Food and Agriculture, U.S. Department of Agriculture, Mclntire Stennis capacity grants # MISZ-069390.
Acknowledgments
This publication is a contribution of the Forest and Wildlife Research Center, Mississippi State University. This material is based upon work that is supported by the National Institution of Food and Agriculture, U.S. Department of Agriculture, Mclntire Stennis capacity grant # MISZ-069390. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the U.S. Department of Agriculture. Special thanks to my fellow colleagues in the College of Forest Resources at Mississippi State University for their endless assistance to this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Groot, R.S.; Wilson, M.A.; Boumans, R.M. A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol. Econ. 2002, 41, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.D.; Arthur, M.A. Implications of a predicted shift from upland oaks to red maple on forest hydrology and nutrient availability. Can. J. For. Res. 2010, 40, 716–726. [Google Scholar] [CrossRef]
- Corti, G.; Agnelli, A.; Cocco, S.; Cardelli, V.; Masse, J.; Courchesne, F. Soil affects throughfall and stemflow under Turkey oak (Quercus cerris L.). Geoderma 2019, 333, 43–56. [Google Scholar] [CrossRef]
- Dibb, J.E.; Talbot, R.W.; Scheuer, E.; Seid, G.; DeBell, L.; Lefer, B.; Ridley, B. Stratospheric influence on the northern North American free troposphere during TOPSE: 7Be as a stratospheric tracer. J. Geophys. Res. Atmos. 2003, 108. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Barker, A.C. An analysis of throughfall and stemflow in mixed oak stands. Water Resour. Res. 1970, 6, 316–323. [Google Scholar] [CrossRef]
- Carlisle, A.; Brown, A.H.F.; White, E.J. The nutrient content of tree stem flow and ground flora litter and leachates in a Sessile oak (Quercus Petraea) woodland. J. Ecol. 1967, 55, 615–627. [Google Scholar] [CrossRef]
- Abrams, M.D. Where has all the white oak gone? BioScience 2003, 53, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.D.; Arthur, M.A. Increasing red maple leaf litter alters decomposition rates and nitrogen cycling in historically oak-dominated forests of the eastern U.S. Ecosystems 2014, 17, 1371–1383. [Google Scholar] [CrossRef]
- Hutchinson, T.F.; Long, R.P.; Ford, R.D.; Sutherland, E.K. Fire history and the establishment of oaks and maples in second-growth forests. Can. J. For. Res. 2008, 38, 1184–1198. [Google Scholar] [CrossRef] [Green Version]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biol. Biochem. 2004, 36, 1443–1451. [Google Scholar] [CrossRef]
- BassiriRad, H.; Lussenhop, J.F.; Sehtiya, H.L.; Borden, K.K. Nitrogen deposition potentially contributes to oak regeneration failure in the Midwestern temperate forests of the USA. Oecologia 2015, 177, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hanberry, B.B.; Abrams, M.D. Does white-tailed deer density affect tree stocking in forests of the Eastern United States? Ecol. Process. 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Cote, S.D.; Rooney, T.P.; Tremblay, J.P.; Dussault, C.; Waller, D.M. Ecological impacts of deer overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Liang, J.; Cumming, J.R.; Lee, E.; Welsh, A.B.; Watson, J.V.; Zhou, M. Fundamental shifts of central hardwood forests under climate change. Ecol. Model. 2016, 332, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Olson, M.G.; Stevenson, A.P.; Knapp, B.O.; Kabrick, J.M.; Jensen, R.G. Is there evidence of mesophication of oak forests in the Missouri Ozarks? In Proceedings of the 19th Central Hardwood Forest Conference, Carbondale, IL, USA, 10–12 March 2014; Groninger, J.W., Holzmueller, E.J., Nielsen, C.K., Dey, D.C., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; pp. 139–153. [Google Scholar]
- Chapman, J.I.; McEwan, R.W. Thirty years of compositional change in an old-growth temperate forest: The role of topographic gradients in oak-maple dynamics. PLoS ONE 2016, 11, e0160238. [Google Scholar] [CrossRef]
- Kreye, J.K.; Varner, J.M.; Hamby, G.W.; Kane, J.M. Mesophytic litter dampens flammability in fire-excluded pyrophytic oak-hickory woodlands. Ecosphere 2018, 9, e02078. [Google Scholar] [CrossRef]
- Palus, J.D.; Goebel, P.C.; Hix, D.M.; Matthews, S.N. Structural and compositional shifts in forests undergoing mesophication in the Wayne National Forest, southeastern Ohio. For. Ecol. Manag. 2018, 430, 413–420. [Google Scholar] [CrossRef]
- Abrams, M.D. Fire and the development of oak forests. BioScience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Adriaenssens, S.; Hansen, K.; Staelens, J.; Wuyts, K.; De Schrijver, A.; Baeten, L.; Boeckx, P.; Samson, R.; Verheyen, K. Throughfall deposition and canopy exchange processes along a vertical gradient within the canopy of beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst). Sci. Total Environ. 2012, 420, 168–182. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Levia, D.F.; Inamdar, S.P.; Lepori-Bui, M.; Mitchell, M.J. The effects of phenoseason and storm characteristics on throughfall solute washoff and leaching dynamics from a temperate deciduous forest canopy. Sci. Total Environ. 2012, 430, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Siegert, C.M.; Drotar, N.A.; Alexander, H.D. Spatial and temporal variability of throughfall among oak and co-occurring non-oak tree species in an upland hardwood forest. Geosciences 2019, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Levia, D.F.; Frost, E.E. Variability of throughfall volume and solute inputs in wooded ecosystems. Prog. Phys. Geogr. 2006, 30, 605–632. [Google Scholar] [CrossRef]
- Levia, D.F.; Germer, S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands. Rev. Geophys. 2015, 53, 673–714. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E.; Flores Laureano, J.S.; Price, A.G. Throughfall and throughfall spatial variability in Madrean oak forest communities of northeastern Mexico. J. Hydrol. 2004, 297, 124–135. [Google Scholar] [CrossRef]
- Bhat, S.; Jacobs, J.M.; Bryant, M.L. The chemical composition of rainfall and throughfall in five forest communities: A case study in Fort Benning, Georgia. Water Air Soil Pollut. 2011, 218, 323–332. [Google Scholar] [CrossRef]
- Bryant, M.L.; Bhat, S.; Jacobs, J.M. Measurements and modeling of throughfall variability for five forest communities in the southeastern US. J. Hydrol. 2005, 312, 95–108. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Jonard, F.; Ponette, Q. Spatial and temporal patterns of throughfall volume in a deciduous mixed-species stand. J. Hydrol. 2011, 400, 244–254. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Siegert, C.M.; Levia, D.F.; Scheick, C.E. Effects of wind-driven rainfall on stemflow generation between codominant tree species with differing crown characteristics. Agric. For. Meteorol. 2011, 151, 1277–1286. [Google Scholar] [CrossRef]
- Siegert, C.M.; Levia, D.F. Seasonal and meteorological effects on differential stemflow funneling ratios for two deciduous tree species. J. Hydrol. 2014, 519, 446–454. [Google Scholar] [CrossRef]
- Duchesne, L.; Ouimet, R.; Camiré, C.; Houle, D. Seasonal nutrient transfers by foliar resorption, leaching, and litter fall in a northern hardwood forest at Lake Clair Watershed, Quebec, Canada. Can. J. For. Res. 2001, 31, 333–344. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Ponette, Q. Effects of biological and meteorological factors on stemflow chemistry within a temperate mixed oak–beech stand. Sci. Total Environ. 2008, 393, 72–83. [Google Scholar] [CrossRef]
- Wuyts, K.; Adriaenssens, S.; Staelens, J.; Wuytack, T.; Van Wittenberghe, S.; Boeckx, P.; Samson, R.; Verheyen, K. Contributing factors in foliar uptake of dissolved inorganic nitrogen at leaf level. Sci. Total Environ. 2015, 505, 992–1002. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Ponette, Q. Spatial and temporal patterns of throughfall chemistry within a temperate mixed oak–beech stand. Sci. Total Environ. 2008, 397, 215–228. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/HomePage.htm (accessed on 15 October 2019).
- Arguez, A.; Durre, I.; Applequist, S.; Squires, M.; Vose, R.; Yin, X.; Bilotta, R. NOAA’s U.S. Climate Normals (1981-2010); National Centers for Environmental Information: Asheville, NC, USA, 2010.
- Keim, R.F.; Skaugset, A.E.; Weiler, M. Temporal persistence of spatial patterns in throughfall. J. Hydrol. 2005, 314, 263–274. [Google Scholar] [CrossRef]
- Robertson, S.M.C.; Hornung, M.; Kennedy, V.H. Water chemistry of throughfall and soil water under four tree species at Gisburn, northwest England, before and after felling. For. Ecol. and Manag. 2000, 129, 101–117. [Google Scholar] [CrossRef]
- Levia, D.F.; Herwitz, S.R. Physical properties of water in relation to stemflow leachate dynamics: Implications for nutrient cycling. Can. J. For. Res. 2000, 30, 662–666. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Marin, C.T.; Bouten, W.; Sevink, J. Gross rainfall and its partitioning into throughfall, stemflow and evaporation of intercepted water in four forest ecosystems in western Amazonia. J. Hydrol. 2000, 237, 40–57. [Google Scholar] [CrossRef]
- Andre, F.; Jonard, M.; Ponette, Q. Influence of species and rain event characteristics on stemflow volume in a temperate mixed oak-beech stand. Hydrol. Process. 2008, 22, 4455–4466. [Google Scholar] [CrossRef]
- Siegle-Gaither, M. Using Deuterium and Oxygen-18 Isotopes to Better Understand Stemflow Generation Mechanisms; Mississippi State University: Starkville, MS, USA, 2017. [Google Scholar]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemflow and interception: Effect of forest type, ground cover and climate. Hydrol. Process. 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Levia, D.F.; Frost, E.E. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems. J. Hydrol. 2003, 274, 1–29. [Google Scholar] [CrossRef]
- Thomas, R.; Bennett, N. Estimating bark thicknesses of common Appalachian hardwoods. In Proceedings of the 19th Central Hardwood Forest Conference, Carbondale, IL, USA, 10–12 March 2014; Groninger, J., Holzmueller, E., Nielsen, C., Dey, D., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 388. [Google Scholar]
- Hengst, G.E.; Dawson, J.O. Bark properties and fire resistance of selected tree species from the central hardwood region of North America. Can. J. For. Res. 1994, 24, 688–696. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Kush, J.S.; Fan, Z. Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire prone ecosystem. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Link, T.E.; Unsworth, M.; Marks, D. The dynamics of rainfall interception by a seasonal temperate rainforest. Agric. For. Meteorol. 2004, 124, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Pypker, T.G.; Unsworth, M.H.; Bond, B.J. The role of epiphytes in rainfall interception by forests in the Pacific Northwest. II. Field measurements at the branch and canopy scale. Can. J. For. Res. 2006, 36, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-P.; Wang, Z.-N.; Berndtsson, R.; Zhang, Y.-F.; Pan, Y.-X. Desert shrub stemflow and its significance in soil moisture replenishment. Hydrol. Earth Syst. Sci. 2011, 15, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Silva, I.C.; Okumura, T. Throughfall, stemflow and interception loss in a mixed white oak forest (Quercus serrata Thunb.). J. For. Res. 1996, 1, 123–129. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K. Seasonal variation in throughfall and stemflow chemistry beneath a European beech (Fagus sylvatica) tree in relation to canopy phenology. Can. J. For. Res. 2007, 37, 1359–1372. [Google Scholar] [CrossRef]
- Hörmann, G.; Branding, A.; Clemen, T.; Herbst, M.; Hinrichs, A.; Thamm, F. Calculation and simulation of wind controlled canopy interception of a beech forest in Northern Germany. Agric. For. Meteorol. 1996, 79, 131–148. [Google Scholar] [CrossRef]
- Verstraeten, A.; Neirynck, J.; Genouw, G.; Cools, N.; Roskams, P.; Hens, M. Impact of declining atmospheric deposition on forest soil solution chemistry in Flanders, Belgium. Atmos. Environ. 2012, 62, 50–63. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Spatial variability and temporal stability of throughfall water under a dominant beech (Fagus sylvatica L.) tree in relationship to canopy cover. J. Hydrol. 2006, 330, 651–662. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Rainfall partitioning into throughfall, stemflow, and interception within a single beech (Fagus sylvatica L.) canopy: Influence of foliation, rain event characteristics, and meteorology. Hydrol. Process. 2008, 22, 33–45. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Pan, Y.; Hu, R. Relative contribution of biotic and abiotic factors to stemflow production and funneling efficiency: A long-term field study on a xerophytic shrub species in Tengger Desert of northern China. Agric. For. Meteorol. 2020, 280, 107781. [Google Scholar] [CrossRef]
- Skau, C.M. Interception, throughfall, and stemflow in Utah and Alligator Juniper cover types of northern Arizona. For. Sci. 1964, 10, 283–287. [Google Scholar]
- Siegert, C.M.; Levia, D.F.; Leathers, D.J.; Van Stan, J.T.; Mitchell, M.J. Do storm synoptic patterns affect biogeochemical fluxes from temperate deciduous forest canopies? Biogeochemistry 2017, 132, 273–292. [Google Scholar] [CrossRef]
- Potter, C.S.; Ragsdale, H.L.; Swank, W.T. Atmospheric deposition and foliar leaching in a regenerating southern Appalachian forest canopy. J. Ecol. 1991, 79, 97–115. [Google Scholar] [CrossRef]
- Tukey, H.B. The leaching of substances from plants. Annu. Rev. Plant Physiol. 1970, 21, 305–324. [Google Scholar] [CrossRef]
- Talkner, U.; Krämer, I.; Hölscher, D.; Beese, F.O. Deposition and canopy exchange processes in central-German beech forests differing in tree species diversity. Plant Soil 2010, 336, 405–420. [Google Scholar] [CrossRef] [Green Version]
- Lovett, G.M.; Nolan, S.S.; Driscoll, C.T.; Fahey, T.J. Factors regulating throughfall flux in a New Hampshire forested landscape. Can. J. For. Res. 1996, 26, 2134–2144. [Google Scholar] [CrossRef]
- Whelan, M.J.; Sanger, L.J.; Baker, M.; Anderson, J.M. Spatial patterns of throughfall and mineral ion deposition in a lowland Norway spruce (Picea abies) plantation at the plot scale. Atmos. Environ. 1998, 32, 3493–3501. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Spatial variability and temporal stability of throughfall deposition under beech (Fagus sylvatica L.) in relationship to canopy structure. Environ. Pollut. 2006, 142, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Lovett, G.M.; Lindberg, S.E. Dry deposition and canopy exchange in a mixed oak forest as determined by analysis of throughfall. J. Appl. Ecol. 1984, 21, 1013–1027. [Google Scholar] [CrossRef]
- Hamburg, S.P.; Lin, T.-C. Throughfall chemistry of an ecotonal forest on the edge of the Great Plains. Can. J. For. Res. 1998, 28, 1456–1463. [Google Scholar] [CrossRef]
- Cape, J.N.; Sheppard, L.J.; Crossley, A.; van Dijk, N.; Tang, Y.S. Experimental field estimation of organic nitrogen formation in tree canopies. Environ. Pollut. 2010, 158, 2926–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Zhao, C.; Xu, W.; Xie, Z. Hydrochemical fluxes in bulk precipitation, throughfall, and stemflow in a mixed evergreen and deciduous broadleaved forest. Forests 2019, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Guerrieri, R.; Vanguelova, E.I.; Michalski, G.; Heaton, T.H.E.; Mencuccini, M. Isotopic evidence for the occurrence of biological nitrification and nitrogen deposition processing in forest canopies. Glob. Chang. Biol. 2015, 21, 4613–4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, R.; Lecha, L.; Mattana, S.; Cáliz, J.; Casamayor, E.O.; Barceló, A.; Michalski, G.; Peñuelas, J.; Avila, A.; Mencuccini, M. Partitioning between atmospheric deposition and canopy microbial nitrification into throughfall nitrate fluxes in a Mediterranean forest. J. Ecol. 2019, 00, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Le Mellec, A.; Meesenburg, H.; Michalzik, B. The importance of canopy-derived dissolved and particulate organic matter (DOM and POM)—Comparing throughfall solution from broadleaved and coniferous forests. Ann. For. Sci. 2010, 67, 411. [Google Scholar] [CrossRef]
- Verstraeten, A.; Verschelde, P.; De Vos, B.; Neirynck, J.; Cools, N.; Roskams, P.; Hens, M.; Louette, G.; Sleutel, S.; De Neve, S. Increasing trends of dissolved organic nitrogen (DON) in temperate forests under recovery from acidification in Flanders, Belgium. Sci. Total Environ. 2016, 553, 107–119. [Google Scholar] [CrossRef]
- Fan, H.B.; Hong, W. Estimation of dry deposition and canopy exchange in Chinese fir plantations. For. Ecol. Manag. 2001, 147, 99–107. [Google Scholar] [CrossRef]
- Moreno, G.; Gallardo, J.F.; Bussotti, F. Canopy modification of atmospheric deposition in oligotrophic Quercus pyrenaica forests of an unpolluted region (central-western Spain). For. Ecol. Manag. 2001, 149, 47–60. [Google Scholar] [CrossRef]
- Mahendrappa, M.K. Chemical composition of stemflow from some eastern Canadian tree species. Can. J. For. Res. 1974, 4, 1–7. [Google Scholar] [CrossRef]
- Chang, S.C.; Matzner, E. The effect of beech stemflow on spatial patterns of soil solution chemistry and seepage fluxes in a mixed beech/oak stand. Hydrol. Process. 2000, 14, 135–144. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Pypker, T.G. A review and evaluation of forest canopy epiphyte roles in the partitioning and chemical alteration of precipitation. Sci. Total Environ. 2015, 536, 813–824. [Google Scholar] [CrossRef]
- Levia, D.F.; Van Stan, J.T.; Siegert, C.M.; Inamdar, S.P.; Mitchell, M.J.; Mage, S.M.; McHale, P.J. Atmospheric deposition and corresponding variability of stemflow chemistry across temporal scales in a mid-Atlantic broadleaved deciduous forest. Atmos. Environ. 2011, 45, 3046–3054. [Google Scholar] [CrossRef]
- Fahey, T.J.; Williams, C.J.; Rooney-Varga, J.N.; Cleveland, C.C.; Postek, K.M.; Smith, S.D.; Bouldin, D.R. Nitrogen deposition in and around an intensive agricultural district in central New York. J. Environ. Qual. 1999, 28, 1585–1600. [Google Scholar] [CrossRef]
- Herrmann, M.; Pust, J.; Pott, R. The chemical composition of throughfall beneath oak, birch and pine canopies in Northwest Germany. Plant Ecol. 2006, 184, 273–285. [Google Scholar] [CrossRef]
- Qualls, R.G.; Haines, B.L. Geochemistry of dissolved organic nutrients in water percolating through a forest ecosystem. Soil Sci. Soc. Am. J. 1991, 55, 1112–1123. [Google Scholar] [CrossRef] [Green Version]
- Augusto, L.; Dupouey, J.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Pitman, R.M.; Vanguelova, E.I.; Benham, S.E. The effects of phytophagous insects on water and soil nutrient concentrations and fluxes through forest stands of the Level II monitoring network in the UK. Sci. Total Environ. 2010, 409, 169–181. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Mack, M.C.; Woods, C.; DeMarco, J.; Treseder, K.K. The influence of tree species on canopy soil nutrient status in a tropical lowland wet forest in Costa Rica. Plant Soil 2009, 318, 47–61. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Van Stan, J.H.; Levia, D.F. Meteorological influences on stemflow generation across diameter size classes of two morphologically distinct deciduous species. Int. J. Biometeorol. 2014, 58, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Stubbins, A.; Silva, L.M.; Dittmar, T.; Van Stan, J.T. Molecular and optical properties of tree-derived dissolved organic matter in throughfall and stemflow from live oaks and eastern red cedar. Front. Earth Sci. 2017, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Fox, J.E.D.; Xu, Z. Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane subtropical moist forest on Ailao Mountains in Yunnan, south-west China. J. Trop. Ecol. 2002, 18, 527–548. [Google Scholar] [CrossRef]
- Lovett, G.M.; Lindberg, S.E. Dry deposition of nitrate to a deciduous forest. Biogeochemistry 1986, 2, 137–148. [Google Scholar] [CrossRef]
- Fowler, D.; Cape, J.N.; Unsworth, M.H.; Mayer, H.; Crowther, J.M.; Jarvis, P.G.; Gardiner, B.; Shuttleworth, W.J. Deposition of atmospheric pollutants on forests. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1989, 324, 247–265. [Google Scholar] [CrossRef]
- Morris, D.M.; Gordon, A.G.; Gordon, A.M. Patterns of canopy interception and throughfall along a topographic sequence for black spruce dominated forest ecosystems in northwestern Ontario. Can. J. For. Res. 2003, 33, 1046–1060. [Google Scholar] [CrossRef]
- Puckett, L.J. Estimates of ion sources in deciduous and coniferous throughfall. Atmos. Environ. 1990, 24, 545–555. [Google Scholar] [CrossRef]
- Hafner, S.D.; Groffman, P.M. Soil nitrogen cycling under litter and coarse woody debris in a mixed forest in New York State. Soil Biol. Biochem. 2005, 37, 2159–2162. [Google Scholar] [CrossRef]
- Neary, A.J.; Gizyn, W.I. Throughfall and stemflow chemistry under deciduous and coniferous forest canopies in South Central Ontario. Can. J. For. Res. 1994, 24, 1089–1100. [Google Scholar] [CrossRef]
- Foster, N.W.; Nicolson, J.A. Acid deposition and nutrient leaching from deciduous vegetation and podzolic soils at the Turkey Lakes watershed. Can. J. Fish. Aquat. Sci. 1988, 45, s96–s100. [Google Scholar] [CrossRef]
- Jo, I.; Fei, S.; Oswalt, C.M.; Domke, G.M.; Phillips, R.P. Shifts in dominant tree mycorrhizal associations in response to anthropogenic impacts. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Throughfall volume per tree basal area (L/m2) of (a) Q. pagoda, (b) Q. shumardii, (c) Q. alba, (d) Q. stellata, (e) C. glabra, and (f) C. ovata during the leafless phase (November–March) and the leafed phase (April–October) in relation to open precipitation.
Figure 1.
Throughfall volume per tree basal area (L/m2) of (a) Q. pagoda, (b) Q. shumardii, (c) Q. alba, (d) Q. stellata, (e) C. glabra, and (f) C. ovata during the leafless phase (November–March) and the leafed phase (April–October) in relation to open precipitation.

Figure 2.
Stemflow volume per tree basal area (L/m2) of (a) Q. pagoda, (b) Q. shumardii, (c) Q. alba, (d) Q. stellata, (e) C. glabra, and (f) C. ovata during the leafless phase (November–March) and the leafed phase (April–October) in relation to open precipitation.
Figure 2.
Stemflow volume per tree basal area (L/m2) of (a) Q. pagoda, (b) Q. shumardii, (c) Q. alba, (d) Q. stellata, (e) C. glabra, and (f) C. ovata during the leafless phase (November–March) and the leafed phase (April–October) in relation to open precipitation.

Figure 3.
Average seasonal solute concentrations in precipitation (PG), throughfall (TF), and stemflow (SF) for (a) dissolved organic carbon, (b) nitrate, (c) ammonium, and (d) with error bars by species.
Figure 3.
Average seasonal solute concentrations in precipitation (PG), throughfall (TF), and stemflow (SF) for (a) dissolved organic carbon, (b) nitrate, (c) ammonium, and (d) with error bars by species.

Figure 4.
Enrichment ratios of (a) throughfall DOC, (b) stemflow DOC, (c) throughfall NO3−, (d) stemflow NO3−, (e) throughfall NH4+, and (f) stemflow NH4+. DON was omitted from analysis due to limited data points. Horizontal dashed line at y = 1 denotes no change in chemistry.
Figure 4.
Enrichment ratios of (a) throughfall DOC, (b) stemflow DOC, (c) throughfall NO3−, (d) stemflow NO3−, (e) throughfall NH4+, and (f) stemflow NH4+. DON was omitted from analysis due to limited data points. Horizontal dashed line at y = 1 denotes no change in chemistry.

Figure 5.
Throughfall enrichment ratios as a function of throughfall depth in centimeters of (a) DOC, (b) NO3−, and (c) NH4+. DON was omitted from analysis due to limited data points.
Figure 5.
Throughfall enrichment ratios as a function of throughfall depth in centimeters of (a) DOC, (b) NO3−, and (c) NH4+. DON was omitted from analysis due to limited data points.

Figure 6.
Stemflow enrichment ratios as a function of stemflow volumetric flux in liters per square meter of tree basal area of (a) DOC, (b) NO3−, and (c) NH4+. DON was omitted from analysis due to limited data points.
Figure 6.
Stemflow enrichment ratios as a function of stemflow volumetric flux in liters per square meter of tree basal area of (a) DOC, (b) NO3−, and (c) NH4+. DON was omitted from analysis due to limited data points.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Average tree characteristics, including diameter at breast height (DBH), crown area, and tree height of the trees, in this study located at Sessums Natural Area. Species codes are used in figures and tables throughout.
Table 1.
Average tree characteristics, including diameter at breast height (DBH), crown area, and tree height of the trees, in this study located at Sessums Natural Area. Species codes are used in figures and tables throughout.
| Tree Species | Species Code | DBH (cm) | Crown Area (m2) | Tree Height (m) | Bark Thickness (cm) | |
|---|---|---|---|---|---|---|
| Shumard oak | Quercus shumardii | QUSH | 65.4 | 47.6 | 40.7 | 0.94 |
| Cherrybark oak | Quercus pagoda | QUPA | 68.6 | 179.1 | 48.0 | 1.07 |
| Post oak | Quercus stellata | QUST | 59.1 | 147.4 | 25.2 | 1.35 |
| White oak | Quercus alba | QUAL | 66.6 | 132.6 | 45.1 | 1.67 |
| Pignut hickory | Carya glabra | CAGL | 43.9 | 76.6 | 23.1 | 1.13 |
| Shagbark hickory | Carya ovata | CAOV | 35.7 | 44.4 | 21.8 | 0.64 |
Table 2.
Average seasonal enrichment ratio of solutes by species. DON (Dissolved Organic Nitrogen) was not included in this table due to limited data during the leafless season.
Table 2.
Average seasonal enrichment ratio of solutes by species. DON (Dissolved Organic Nitrogen) was not included in this table due to limited data during the leafless season.
| Leafed | Leafless | |||||
|---|---|---|---|---|---|---|
| DOC | NO3− | NH4+ | DOC | NO3− | NH4+ | |
| THROUGHFALL | ||||||
| Cherrybark Oak | 3.66 | 0.59 | 0.93 | 1.68 | 0.88 | 0.57 |
| Shumard Oak | 3.00 | 0.35 | 0.85 | 2.19 | 0.88 | 0.59 |
| White Oak | 5.17 | 1.27 | 1.52 | 1.55 | 3.33 | 0.10 |
| Post Oak | 5.65 | 1.32 | 1.29 | 2.35 | 2.94 | 0.14 |
| Pignut Hickory | 2.87 | 1.52 | 1.22 | 4.12 | 33.82 | 0.03 |
| Shagbark Hickory | 4.36 | 1.29 | 0.28 | 2.10 | 2.50 | 0.04 |
| Average | 4.18 | 0.89 | 1.00 | 2.31 | 1.99 | 0.75 |
| STEMFLOW | ||||||
| Cherrybark Oak | 6.07 | 0.81 | 0.26 | 14.03 | 4.32 | 2.78 |
| Shumard Oak | 15.32 | 2.20 | 0.49 | 13.55 | 0.66 | 5.72 |
| White Oak | 10.74 | 0.50 | 0.26 | 16.18 | 0.67 | 0.08 |
| Post Oak | 17.87 | 3.48 | 0.76 | 18.70 | 2.27 | 1.61 |
| Pignut Hickory | 10.96 | 3.52 | 0.90 | 32.67 | 1.27 | 0.70 |
| Shagbark Hickory | 122.12 | 24.56 | 7.35 | 61.06 | 7.26 | 19.86 |
| Average | 32.02 | 6.40 | 2.07 | 24.33 | 2.88 | 5.65 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Limpert, K.; Siegert, C. Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest. Forests 2019, 10, 1121. https://doi.org/10.3390/f10121121
AMA Style
Limpert K, Siegert C. Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest. Forests. 2019; 10(12):1121. https://doi.org/10.3390/f10121121
Chicago/Turabian StyleLimpert, Katy, and Courtney Siegert. 2019. "Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest" Forests 10, no. 12: 1121. https://doi.org/10.3390/f10121121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.