On the Biological and Genetic Diversity in Neospora caninum

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Isolates | Source | Country | Reference |
|---|---|---|---|
| NC1 | Brain of congenitally infected dog | United States | [13] |
| NC-2 | Muscle biopsy of naturally infected dog | United States | [16] |
| NC-3 | Brain and spinal cord of dog | United States | [17] |
| NC-4 & NC-5 | Brain of a congenitally dog | United States | [18] |
| NC-6, 7 & 8 | Brain and striated muscle of puppy | United States | [19] |
| NC-9 | Brain of naturally infected puppy | United States | [20] |
| CN1 | Brain and spinal cord of a congenitally infected dog | United States | [9] |
| NC-Liverpool | Cerebrum of congenitally infected puppy | United Kingdom | [21,22] |
| NC-Bahia | Brain of naturally infected adult dog | Brazil | [23] |
| NC-6 Argentina | Oocysts from naturally infected dog | Argentina | [24] |
| WA-K9 | Skin lesions of naturally infected dog | Australia | [25] |
| CZ-4 * | Oocysts from naturally infected dog | Czech Republic | [26] |
| NC-GER1 | Brain and spinal cord of congenitally infected puppy | Germany | [27] |
| Hammondia heydorni-Berlin-1996 | Oocysts from naturally infected dog | Germany | [28] |
| NC-GER2, 3, 4, 5 and NC-GER-6 | Oocystes from naturally infected dog | Germany | [29] |
| NC-GER7, 8 and NC-GER-9 | Oocysts from naturally infected dog | Germany | [30] |
| NC-P1 | Oocysts from naturally infected dog | Portugal | [31] |
| BPA1 & BPA2 | Brain of aborted bovine foetuses | United States | [14] |
| BPA3 & BPA4 | Brain of congenitally infected calf | United States | [32] |
| BPA6 | Brain and/or spinal cord of an aborted bovine foetus | United States | [33] |
| NC-Beef | Naturally infected calf | United States | [34] |
| NC-Illinois | Brain of naturally infected calf | United States | [35] |
| VMDL1 | Brain of aborted beef calf | United States | [36] |
| NC-LivB1 | Brain of stillborn calf | United Kingdom | [37,38] |
| NC-LivB2 | Brain of aborted bovine foetus | United Kingdom | [39] |
| NC-Porto1 | Brain of aborted bovine foetus | Portugal | [40] |
| NC-Sp1 | Brain of aborted bovine foetus | Spain | [41] |
| Five isolates | Brain of four aborted bovine foetuses and one stillborn calf | Spain | [42] |
| NC-Spain 1H | Brain of naturally infected and healthy calf | Spain | [43] |
| NC-Spain 2H, 3H, 4H and 5H, NC-Spain6, 7, 8, 9, 10 | Brain of naturally infected and healthy calf | Spain | [44] |
| JAP1 | Brain and spinal cord of congenitally infected calf | Japan | [45,46] |
| BT-2 & JAP-2 JAP-5 JAP-4 | Brain of congenitally infected calf Brain and spinal cord of congenitally infected calf Brain of stillborn calf | Japan | [47] |
| BT-3 | Brain of an infected adult cow | Japan | [48] |
| NC-Sheep+ | Brain of naturally infected pregnant sheep | Japan | [49] |
| NC-SweB1 | Brain of stillborn calf | Sweden | [50] |
| NC-VP1 | Brain of a congenitally infected calf | Italy | [51] |
| NCPG1 | Brain and placenta cotyledonary villi of clinically naturally infected calf | Italy | [52] |
| NC-MalB1 | Brain of congenitally infected calf | Malaysia | [53] |
| KBA-1 | Brain of a congenitally infected calf | South Korea | [54] |
| KBA-2 | Brain of aborted bovine foetus | ||
| NC-Kr2 | Brain of naturally infected cow | South Korea | Jeong et al. unpublished |
| BNC-PRI | Brain of congenitally blind calf | Brazil | [55] |
| BCN-PR3 | Brain of aborted bovine foetus | Brazil | [56] |
| No name | Brain of naturally infected sheep | Brazil | [57] |
| Nc-Goiás 1 | Brain of clinically healthy calf | Brazil | [58] |
| NC-Nowra | Brain and spinal cord of congenitally infected calf | Australia | [59] |
| NcNZ1 NcNZ2 NcNZ3 | Brain of naturally infected cow Brain of infected 2-days old calf Brain of stillborn calf | New Zealand | [60] |
| NcIs491 NcIs580 | Brain of aborted bovine foetus | Israel | [61] |
| NC-PolB1 | Brain of naturally infected calf | Poland | [62,63] |
| NcBrBuf-1, 2, 3, 4, 5 | Brain of naturally infected water buffalo | Brazil | [64] |
| Nc-Iran1 | Brain of aborted bovine foetus | Iran | Salehi et al. unpublished |
| NC-deer1 * | Brain of naturally infected white-tailed deer ( Odocoileus virginianus ) | United States | [65] |
| NC-WTDVA-1 NC-WTDVA-2* and 3* | Brain of naturally infected white-tailed deer ( Odocoileus virginianus) | United States | [66] |
2. Comparison of Biological Characteristics between Neospora caninum Isolates
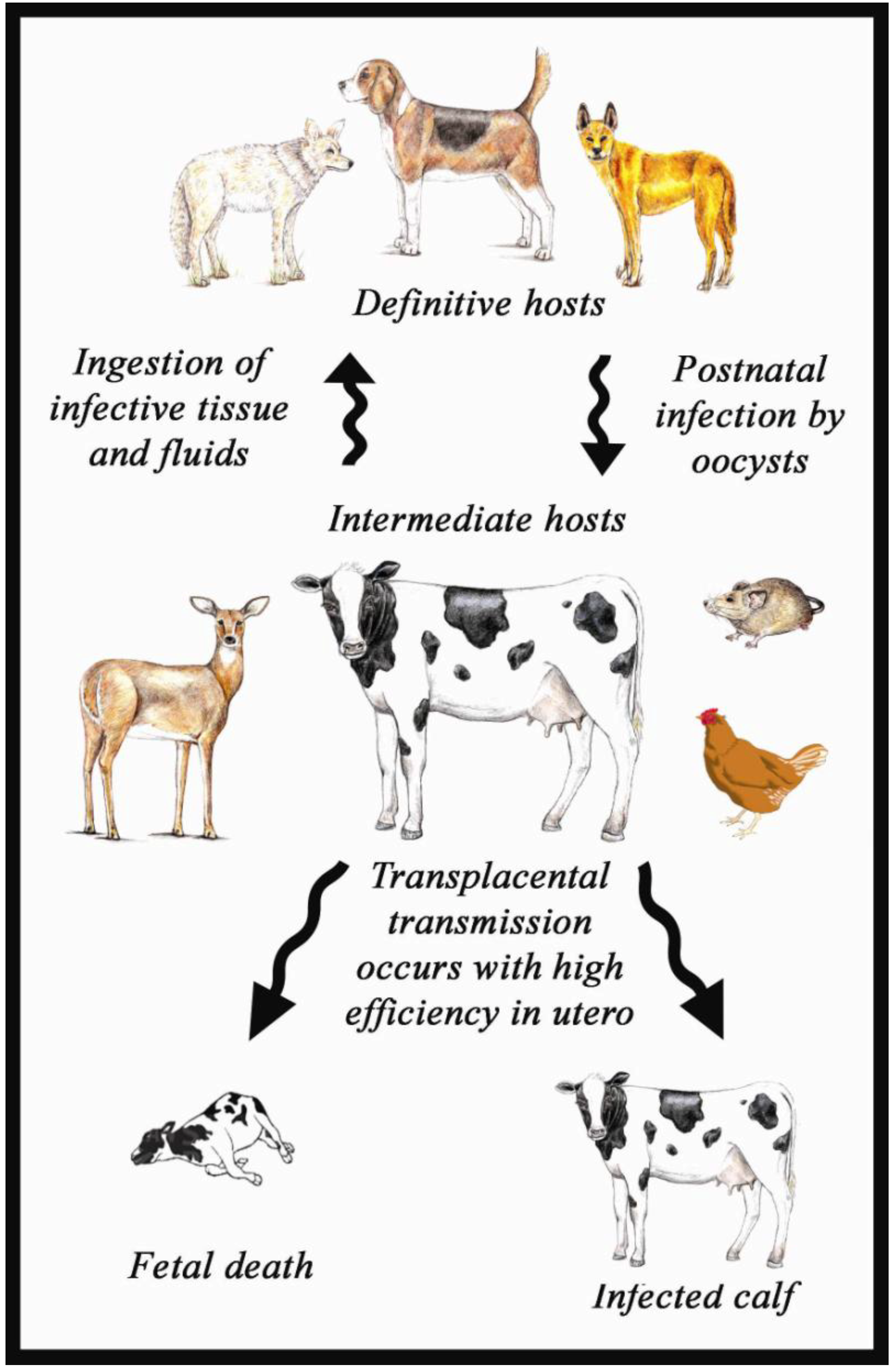
2.1. Life Cycle of N caninum

2.2. Life Cycle Stages and Ultrastructure
2.2.1. Tachyzoite size
| Isolate name | Tachyzoite size (µm) | References |
|---|---|---|
| NC1 | 4.3–4.6 × 2.1–2.3, 4.8–5.3 × 1.8–2.3 | [1] |
| 5.1–8.4 × 1.5–2.5 | [77] | |
| NC-SweB1 | 5 × 1.3 | [50] |
| KBA1 and KBA2 | 5–7 × 1.5–2 | [54] |
| NC-Liverpool | 6 × 2 | [22] |
| BPA1 and BPA2 | 6–8 × 1–2 | [14] |
| JAP1 | 5–6 × 1.5–2 | [46] |
| NC-Sheep+ | 7 × 3 | [49] |
2.2.2. Rhoptries
2.2.3. Micronemes
2.2.4. Tissue cysts
| Source | Tissue cysts size (µm)* | Thickness of the tissue cysts (µm)* | References |
|---|---|---|---|
| Brain of congenitally infected dog | 55 × 25, 65–45, 100 × 77 and 107–75 | 2–3 1.5–3.5 | [1] |
| ND | >1 | [86] | |
| 14–65 long | 0.7–4.5 | [19] | |
| 10–42 × 10–40 | ~ 1 | [18] | |
| Brain of naturally infected dog | 18.1–27.4 × 14.1–23.7 | 0.74–1.12 | [81] |
| Brain of infected Swiss Webster mice | ND | 0.5–4 | [82] |
| Brain of infected bovine foetuses | 6.8–22.7 × 7.4–23.7 | 0.34–1.05, 0.23–0.77, 0.43–1.05 and 0.77–1.22 | [87] |
| 8–10 8–13 | <1 1–2 | [14] | |
| 50 | 2 | [88] | |
| Spinal cord of naturally infected calf | 20–48 14–20 | 1–2 0.53–3.09 and 0.53–1.69 | [83] |
| Brain of naturally infected caprine foetuses | 10–32 15 14–21.8 | 1–2 1 0.51–1.75 | [89] |
| Heart muscle of naturally infected caprine foetus | 10–22 | 1–1.5 | |
| Cerebrum of an aborted goat foetus | 6–20 (mostly 10 µm) | 0.5–1.0 | [90] |
| Brain of gerbil inoculated with oocysts | 24.47–47.07 × 25.33–53.91 | 1.71–2.66 | [57] |
| 28 × 26 and 32 × 27 | >1 | [91] | |
| Brain of mice infected with tachyzoites | 15–107 long | ND | [92] |
| Brain of naturally infected horse | ND | 1.5–3 | [93] |
| ND | 2 | [94] |
2.2.5. Oocysts
| Sources | Size of oocysts (µm)* | Oocyst Number* | References |
|---|---|---|---|
| Dog fed infected mice carcasses | 10–11 | few | [34] |
| ND | 4.5 × 106 from dog1 but few from dog 2 | [67] | |
| Dog fed infected mouse brains | 10.6–12.4 × 10.6–12 | ND | [76] |
| Dog fed infected mouse carcasses | ≤11.5 | 100–29,900 | [35] |
| Dog fed experimentally infected calf tissues | 0–503,300 | ||
| Dog fed experimentally infected calf tissues | ND | 792,000 | [95] |
| Dog fed naturally infected bovine placenta | 10–11 | few | [75] |
| Dog fed muscles of experimentally infected sheep | 10.5–12.2, 10.5–11.7, 9.4–13.4 and 10.2–11.4 | 15 × 105 | [28] |
| Dog fed muscles of experimentally infected goat | 8 × 105 | ||
| Dog fed infected guinea pig | 1 × 106–2 × 106 | ||
| Dog fed brain of naturally infected white tailed deer | 10 | 12,300 | [65] |
| Dog fed brain of naturally infected water buffaloes | ND | 43,500–820,655 | [64] |
| Dog fed brain of naturally infected sheep | 6.56–10.84 × 9.73–11.86 | 27,600 | [57] |
| Naturally infected dog | 10.7 | 19–114,000 / g of faeces | [29] |
| 9.89 × 9.95 | ND | [30] | |
| 10 –13 × 10–11 | one million | [26] | |
| 10.4–12.9 × 10.6–11.8 | 400 / g of faeces | [97] | |
| 9.71–10.2 × 9.19–9.76 | 19–143,000 | [24] | |
| Naturally infected fox and coyote | 10–11 | few | [98] |
| Coyote fed experimentally infected calf tissues | 10 | 500 | [68] |
| Dingo fed experimentally infected calf tissues | 10 × 12 | 1,810 | [69] |
2.3. Pathogenicity
2.3.1. Studies with mice
2.3.2. Studies with gerbils
2.3.3. Studies with cattle
2.3.4. Studies with sheep
2.4. Antigenicity and Proteins
2.5. DNA Analyses
2.5.1. 18S-like ribosomal DNA (small subunit-rDNA)
2.5.2. Internal transcribed spacer sequences (ITS1)
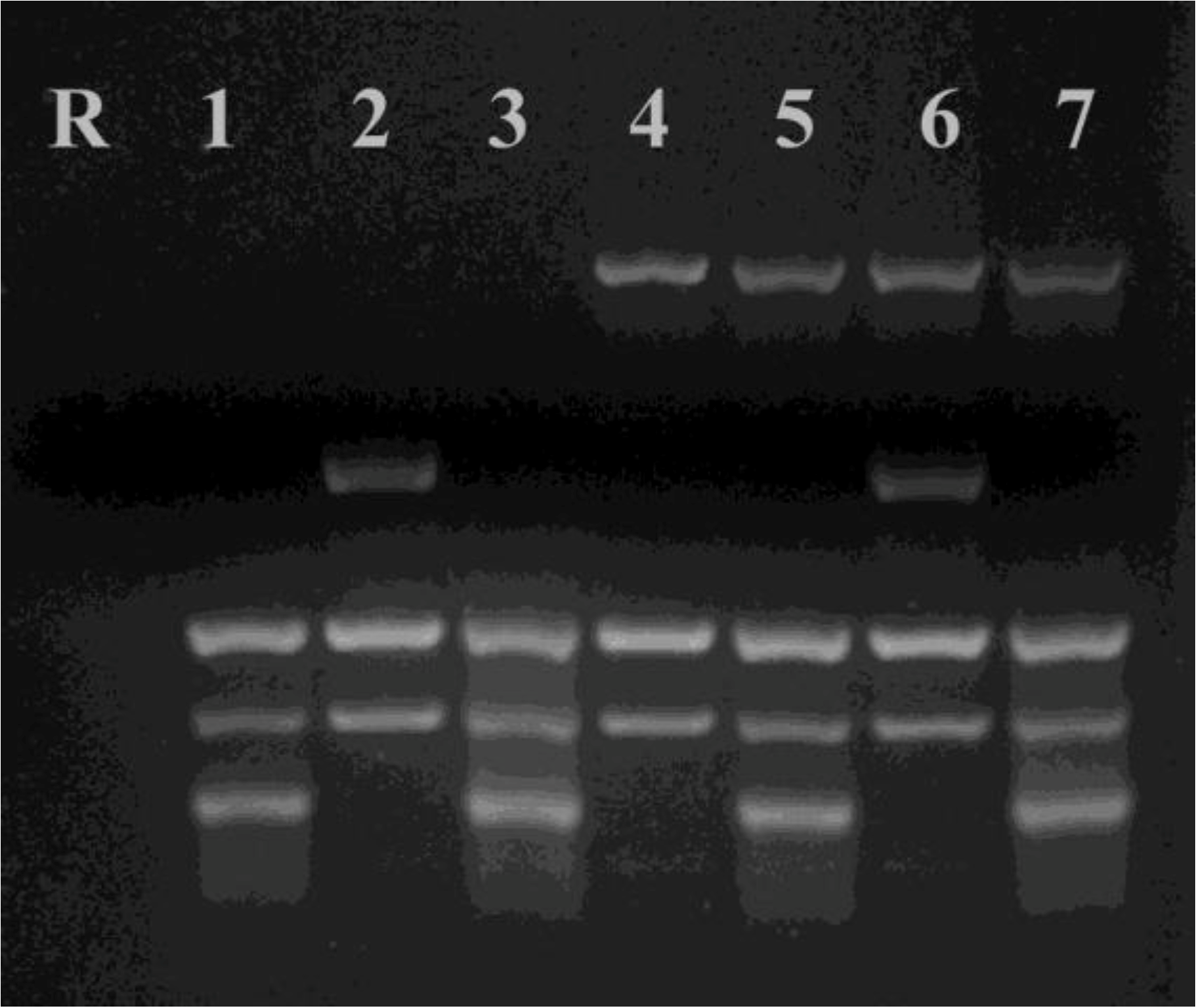
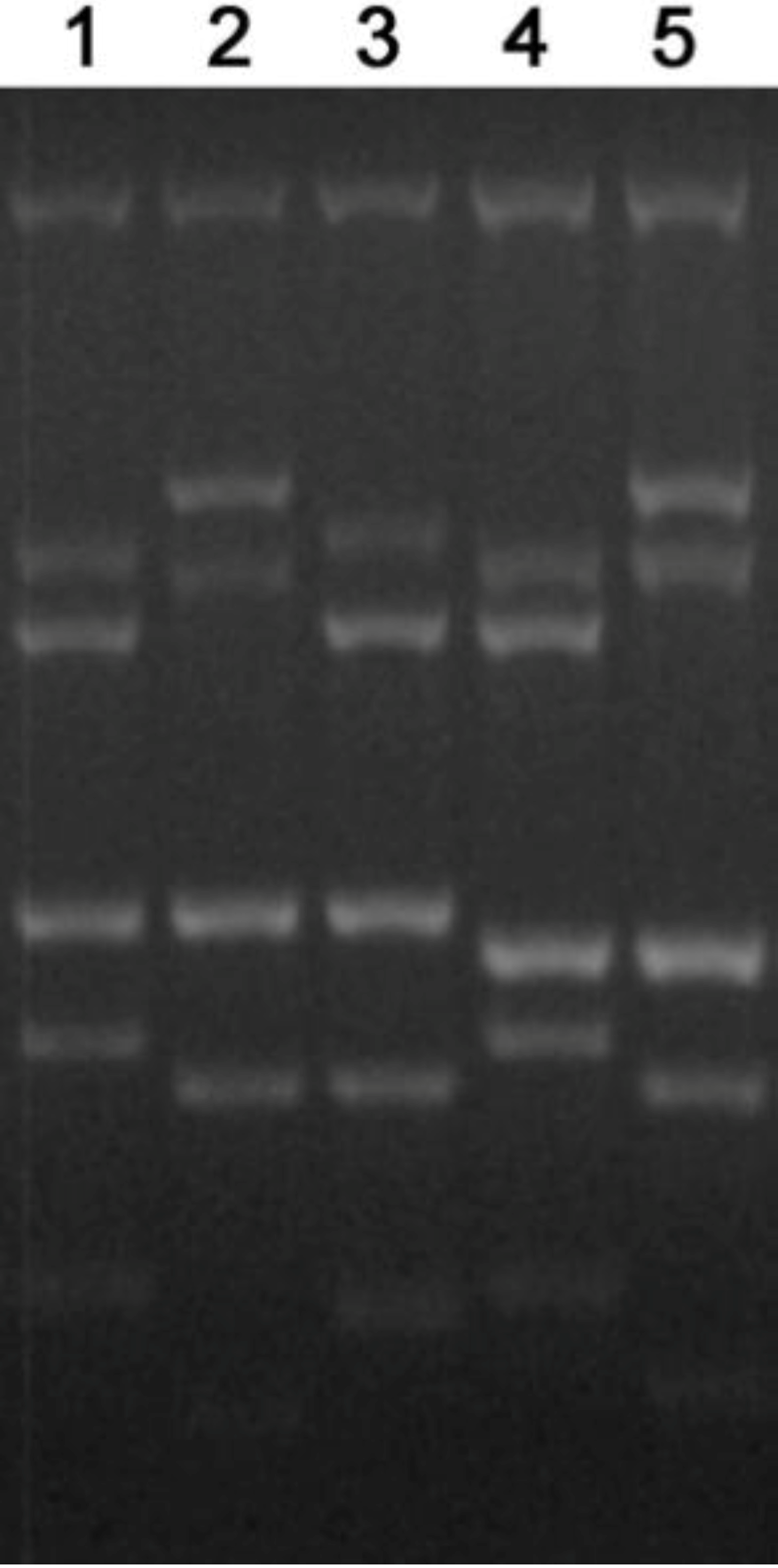
2.5.3. RAPD-PCR
2.5.4. Nc5 repeat
2.5.5. Mini- and microsatellites


2.5.6. Protein-encoding genes
2.6. Growth Rate
3. Discussion
Acknowledgements
References and Notes
- Dubey, J.P.; Carpenter, J.L.; Speer, C.A.; Topper, M.J.; Uggla, A. Newly recognized fatal protozoan disease of dogs. J. Amer. Vet. Med. Assn. 1988, 192, 1269–1285. [Google Scholar]
- Dubey, J.P. Review of Neospora caninum and neosporosis in animals. Korean J. Parasitol. 2003, 41, 1–16. [Google Scholar] [CrossRef]
- Reichel, M.P.; Ellis, J.T.; Dubey, J.P. Neosporosis and hammondiosis in dogs. J. Small Anim. Pract. 2007, 48, 308–312. [Google Scholar] [CrossRef]
- Bjerkås, I.; Dubey, J.P. Evidence that Neospora caninum is identical to the Toxoplasma-like parasite of Norwegian dogs. Acta. Vet. Scand. 1991, 32, 407–410. [Google Scholar]
- Bjerkås, I.; Mohn, S.F.; Presthus, J. Unidentified cyst-forming sporozoon causing encephalomyelitis and myositis in dogs. Z. Parasitenkd 1984, 70, 271–274. [Google Scholar] [CrossRef]
- Thilsted, J.P.; Dubey, J.P. Neosporosis-like abortions in a herd of dairy cattle. J. Vet. Diagn. Invest. 1989, 1, 205–209. [Google Scholar] [CrossRef]
- Dubey, J.P. Neosporosis - a newly recognised protozoal infection. Comp. Pathol. Bull. 1992, 24, 4–6. [Google Scholar]
- Dubey, J.P.; Lindsay, D.S. Neosporosis in dogs. Vet. Parasitol. 1990, 36, 147–151. [Google Scholar] [CrossRef]
- Marsh, A.E.; Barr, B.C.; Packham, A.E.; Conrad, P.A. Description of a new Neospora species (Protozoa: Apicomplexa: Sarcocystidae). J. Parasitol. 1998, 84, 983–991. [Google Scholar] [CrossRef]
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 2007, 20, 323–367. [Google Scholar] [CrossRef]
- Larson, R.L.; Hardin, D.K.; Pierce, V.L. Economic considerations for diagnostic and control options for Neospora caninum-induced abortions in endemically infected herds of beef cattle. J. Amer. Vet. Med. Assn. 2004, 224, 1597–1604. [Google Scholar] [CrossRef]
- Reichel, M.P.; Ellis, J.T. If control of Neospora caninum infection is technically feasible does it make economic sense? Vet. Parasitol. 2006, 142, 23–34. [Google Scholar] [CrossRef]
- Dubey, J.P.; Hattel, A.L.; Lindsay, D.S.; Topper, M.J. Neonatal Neospora caninum infection in dogs: isolation of the causative agent and experimental transmission. J. Amer. Vet. Med. Assn. 1988, 193, 1259–1263. [Google Scholar]
- Conrad, P.A.; Barr, B.C.; Sverlow, K.W.; Anderson, M.; Daft, B.; Kinde, H.; Dubey, J.P.; Munson, L.; Ardans, A. In vitro isolation and characterization of a Neospora sp. from aborted bovine foetuses. Parasitology 1993, 106, 239–249. [Google Scholar] [CrossRef]
- Schock, A.; Innes, E.A.; Yamane, I.; Latham, S.M.; Wastling, J.M. Genetic and biological diversity among isolates of Neospora caninum. Parasitology 2001, 123, 13–23. [Google Scholar]
- Hay, W.H.; Shell, L.G.; Lindsay, D.S.; Dubey, J.P. Diagnosis and treatment of Neospora caninum infection in a dog. J. Amer. Vet. Med. Assn. 1990, 197, 87–89. [Google Scholar]
- Cuddon, P.; Lin, D.S.; Bowman, D.D.; Lindsay, D.S.; Miller, T.K.; Duncan, I.D.; deLahunta, A.; Cummings, J.; Suter, M.; Cooper, B. Neospora caninum infection in English Springer Spaniel littermates. Diagnostic evaluation and organism isolation. J. Vet. Intern. Med. 1992, 6, 325–332. [Google Scholar] [CrossRef]
- Dubey, J.P.; Dorough, K.R.; Jenkins, M.C.; Liddell, S.; Speer, C.A.; Kwok, O.C.; Shen, S.K. Canine neosporosis: clinical signs, diagnosis, treatment and isolation of Neospora caninum in mice and cell culture. Int. J. Parasitol. 1998, 28, 1293–1304. [Google Scholar] [CrossRef]
- Dubey, J.P.; Sreekumar, C.; Knickman, E.; Miska, K.B.; Vianna, M.C.; Kwok, O.C.; Hill, D.E.; Jenkins, M.C.; Lindsay, D.S.; Greene, C.E. Biologic, morphologic, and molecular characterisation of Neospora caninum isolates from littermate dogs. Int. J. Parasitol. 2004, 34, 1157–1167. [Google Scholar] [CrossRef]
- Dubey, J.P.; Vianna, M.C.; Kwok, O.C.; Hill, D.E.; Miska, K.B.; Tuo, W.; Velmurugan, G.V.; Conors, M.; Jenkins, M.C. Neosporosis in Beagle dogs: clinical signs, diagnosis, treatment, isolation and genetic characterization of Neospora caninum. Vet. Parasitol. 2007, 149, 158–166. [Google Scholar] [CrossRef]
- Barber, J.; Trees, A.J.; Owen, M. Isolation of Neospora caninum from a British Dog. Vet. Record 1993, 133, 531–532. [Google Scholar]
- Barber, J.S.; Holmdahl, O.J.M.; Owen, M.R.; Guy, F.; Uggla, A.; Trees, A.J. Characterization of the first European isolate of Neospora caninum (Dubey, Carpenter, Speer, Topper and Uggla). Parasitology 1995, 111, 563–568. [Google Scholar] [CrossRef]
- Gondim, L.F.; Pinheiro, A.M.; Santos, P.O.; Jesus, E.E.; Ribeiro, M.B.; Fernandes, H.S.; Almeida, M.A.; Freire, S.M.; Meyer, R.; McAllister, M.M. Isolation of Neospora caninum from the brain of a naturally infected dog, and production of encysted bradyzoites in gerbils. Vet. Parasitol. 2001, 101, 1–7. [Google Scholar] [CrossRef]
- Basso, W.; Venturini, L.; Venturini, M.C.; Hill, D.E.; Kwok, O.C.; Shen, S.K.; Dubey, J.P. First isolation of Neospora caninum from the feces of a naturally infected dog. J. Parasitol. 2001, 87, 612–618. [Google Scholar] [CrossRef]
- McInnes, L.M.; Irwin, P.; Palmer, D.G.; Ryan, U.M. In vitro isolation and characterisation of the first canine Neospora caninum isolate in Australia. Vet. Parasitol. 2006, 137, 355–363. [Google Scholar] [CrossRef]
- Šlapeta, J.R.; Modrý, D.; Kyselová, I.; Horejš, R.; Lukeš, J.; Koudela, B. Dog shedding oocysts of Neospora caninum: PCR diagnosis and molecular phylogenetic approach. Vet. Parasitol. 2002, 109, 157–167. [Google Scholar] [CrossRef]
- Peters, M.; Wagner, F.; Schares, G. Canine neosporosis: clinical and pathological findings and first isolation of Neospora caninum in Germany. Parasitol. Res. 2000, 86, 1–7. [Google Scholar] [CrossRef]
- Schares, G.; Heydorn, A.O.; Cüppers, A.; Conraths, F.J.; Mehlhorn, H. Hammondia heydorni-like oocysts shed by a naturally infected dog and Neospora caninum NC-1 cannot be distinguished. Parasitol. Res. 2001, 87, 808–816. [Google Scholar] [CrossRef]
- Schares, G.; Pantchev, N.; Barutzki, D.; Heydorn, A.O.; Bauer, C.; Conraths, F.J. Oocysts of Neospora caninum, Hammondia heydorni, Toxoplasma gondii and Hammondia hammondi in faeces collected from dogs in Germany. Int. J. Parasitol. 2005, 35, 1525–1537. [Google Scholar] [CrossRef]
- Basso, W.; Schares, S.; Barwald, A.; Herrmann, D.C.; Conraths, F.J.; Pantchev, N.; Vrhovec, M.G.; Schares, G. Molecular comparison of Neospora caninum oocyst isolates from naturally infected dogs with cell culture-derived tachyzoites of the same isolates using nested polymerase chain reaction to amplify microsatellite markers. Vet. Parasitol. 2009, 160, 43–50. [Google Scholar] [CrossRef]
- Basso, W.; Herrmann, D.C.; Conraths, F.J.; Pantchev, N.; Vrhovec, M.G.; Schares, G. First isolation of Neospora caninum from the faeces of a dog from Portugal. Vet. Parasitol. 2009, 159, 162–166. [Google Scholar] [CrossRef]
- Barr, B.C.; Rowe, J.D.; Sverlow, K.W.; BonDurant, R.H.; Ardans, A.A.; Oliver, M.N.; Conrad, P.A. Experimental reproduction of bovine fetal Neospora infection and death with a bovine Neospora isolate. J. Vet. Diagn. Invest. 1994, 6, 207–215. [Google Scholar] [CrossRef]
- Conrad, P.A.; Barr, B.C.; Anderson, M.L.; Sverlow, K.L. Recombinant neospora antigens and their uses. U.S. Patent 6,716,423, 2004. [Google Scholar]
- McAllister, M.M.; Dubey, J.P.; Lindsay, D.S.; Jolley, W.R.; Wills, R.A.; McGuire, A.M. Dogs are definitive hosts of Neospora caninum. Int. J. Parasitol. 1998, 28, 1473–1478. [Google Scholar] [CrossRef]
- Gondim, L.F.; Gao, L.; McAllister, M.M. Improved production of Neospora caninum oocysts, cyclical oral transmission between dogs and cattle, and in vitro isolation from oocysts. J. Parasitol. 2002, 88, 1159–1163. [Google Scholar] [CrossRef]
- Hyun, C.; Gupta, G.D.; Marsh, A.E. Sequence comparison of Sarcocystis neurona surface antigen from multiple isolates. Vet. Parasitol. 2003, 112, 11–20. [Google Scholar] [CrossRef]
- Davison, H.C.; Trees, A.J.; Guy, F.; Otter, A.; Holt, J.J.; Simpson, V.R.; Jeffrey, M. Isolation of bovine Neospora in Britain. Vet. Record 1997, 141, 607. [Google Scholar]
- Davison, H.C.; Guy, F.; Trees, A.J.; Ryce, C.; Ellis, J.T.; Otter, A.; Jeffrey, M.; Simpson, V.R.; Holt, J.J. In vitro isolation of Neospora caninum from a stillborn calf in the UK. Res. Vet. Sci. 1999, 67, 103–105. [Google Scholar] [CrossRef]
- Trees, A.J.; Williams, D.J.L. Neosporosis in the United Kingdom. Int. J. Parasitol. 2000, 30, 891–893. [Google Scholar]
- Canada, N.; Meireles, C.; Rocha, A.; Sousa, S.; Thompson, G.; Dubey, J.P.; Romand, S.; Thulliez, P.; Correia da Costa, J.M. First Portuguese isolate of Neospora caninum from an aborted fetus from a dairy herd with endemic neosporosis. Vet. Parasitol. 2002, 110, 11–15. [Google Scholar] [CrossRef]
- Canada, N.; Meireles, C.S.; Mezo, M.; González-Warleta, M.; Correia da Costa, J.M.; Sreekumar, C.; Hill, D.E.; Miska, K.B.; Dubey, J.P. First isolation of Neospora caninum from an aborted bovine fetus in Spain. J. Parasitol. 2004, 90, 863–864. [Google Scholar] [CrossRef]
- González-Warleta, M.; Castro-Hermida, J.A.; Carro-Corral, C.; Cortizo-Mella, J.; Mezo, M. Epidemiology of neosporosis in dairy cattle in Galicia (NW Spain). Parasitol. Res. 2008, 102, 243–249. [Google Scholar]
- Rojo-Montejo, S.; Collantes-Fernandez, E.; Regidor-Cerrillo, J.; Alvarez-Garcia, G.; Marugan-Hernandez, V.; Pedraza-Diaz, S.; Blanco-Murcia, J.; Prenafeta, A.; Ortega-Mora, L.M. Isolation and characterization of a bovine isolate of Neospora caninum with low virulence. Vet. Parasitol. 2009, 159, 7–16. [Google Scholar] [CrossRef]
- Regidor-Cerrillo, J.; Gómez-Bautista, M.; Pereira-Bueno, J.; Aduriz, G.; Navarro-Lozano, V.; Risco-Castillo, V.; Férnandez-García, A.; Pedraza-Díaz, S.; Ortega-Mora, L.M. Isolation and genetic characterization of Neospora caninum from asymptomatic calves in Spain. Parasitology 2008, 135, 1651–1659. [Google Scholar] [CrossRef]
- Yamane, I.; Kokuho, T.; Shimura, K.; Eto, M.; Haritani, M.; Ouchi, Y.; Sverlow, K.W.; Conrad, P.A. In vitro isolation of a bovine Neospora in Japan. Vet. Record 1996, 138, 652–652. [Google Scholar]
- Yamane, I.; Kokuho, T.; Shimura, K.; Eto, M.; Shibahara, T.; Haritani, M.; Ouchi, Y.; Sverlow, K.; Conrad, P.A. In vitro isolation and characterisation of a bovine Neospora species in Japan. Res. Vet. Sci. 1997, 63, 77–80. [Google Scholar] [CrossRef]
- Yamane, I.; Shibahara, T.; Kokuho, T.; Shimura, K.; Hamaoka, T.; Haritani, M.; Conrad, P.A.; Park, C.H.; Sawada, M.; Umemura, T. An improved isolation technique for bovine Neospora species. J. Vet. Diagn. Invest. 1998, 10, 364–368. [Google Scholar] [CrossRef]
- Sawada, M.; Kondo, H.; Tomioka, Y.; Park, C.; Morita, T.; Shimada, A.; Umemura, T. Isolation of Neospora caninum from the brain of a naturally infected adult dairy cow. Vet. Parasitol. 2000, 90, 247–252. [Google Scholar] [CrossRef]
- Koyama, T.; Kobayashi, Y.; Omata, Y.; Yamada, M.; Furuoka, H.; Maeda, R.; Matsui, T.; Saito, A.; Mikami, T. Isolation of Neospora caninum from the brain of a pregnant sheep. J. Parasitol. 2001, 87, 1486–1488. [Google Scholar] [CrossRef]
- Stenlund, S.; Bjorkman, C.; Holmdahl, O.J.; Kindahl, H.; Uggla, A. Characterization of a Swedish bovine isolate of Neospora caninum. Parasitol. Res. 1997, 83, 214–219. [Google Scholar] [CrossRef]
- Magnino, S.; Vigo, P.G.; Fabbi, M.; Colombo, M.; Bandi, C.; Genchi, C. Isolation of a bovine Neospora from a newborn calf in Italy. Vet. Record 1999, 144, 456. [Google Scholar]
- Fioretti, D.P.; Rosignoli, L.; Ricci, G.; Moretti, A.; Pasquali, P.; Polidori, G.A. Neospora caninum infection in a clinically healthy calf: Parasitological study and serological follow-up. J. Vet. Med. B–Infect. Dis. Vet. Pub. Health 2000, 47, 47–53. [Google Scholar] [CrossRef]
- Cheah, T.S.; Mattsson, J.G.; Zaini, M.; Sani, R.A.; Jakubek, E.B.; Uggla, A.; Chandrawathani, P. Isolation of Neospora caninum from a calf in Malaysia. Vet. Parasitol. 2004, 126, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Sohn, H.J.; Hwang, W.S.; Hwang, E.K.; Jean, Y.H.; Yamane, I.; Kim, D.Y. In vitro isolation and characterization of bovine Neospora caninum in Korea. Vet. Parasitol. 2000, 90, 147–154. [Google Scholar] [CrossRef]
- Locatelli-Dittrich, R.; Richartz, R.R.T.B.; Joineau, M.E.G.; Pinckney, R.D.; De Sousa, R.S.; Leite, L.C.; Thomaz-Soccol, V. Isolation of Neospora caninum from a blind calf in Parana, southern Brazil. Vet. Record 2003, 153, 366–367. [Google Scholar] [CrossRef]
- Locatelli-Dittrich, R.; Thomaz-Soccol, V.; Richartz, R.R.T.B.; Gasino-Joineau, M.E.; Vander Vinne, R.; Pinckney, R.D. Isolation of Neospora caninum from bovine fetus from a dairy herd in Paraná. Rev. Bras. Parasito. Vet. 2004, 13, 103–109. [Google Scholar]
- Pena, H.F.; Soares, R.M.; Ragozo, A.M.; Monteiro, R.M.; Yai, L.E.; Nishi, S.M.; Gennari, S.M. Isolation and molecular detection of Neospora caninum from naturally infected sheep from Brazil. Vet. Parasitol. 2007, 147, 61–66. [Google Scholar] [CrossRef]
- García-Melo, D.P.; Regidor-Cerrillo, J.; Ortega-Mora, L.M.; Collantes-Fernández, E.; de Oliveira, V.S.F.; de Oliveira, M.A.P.; da Silva, A.C. Isolation and biological characterisation of a new isolate of Neospora caninum from an asymptomatic calf in Brazil. Acta Parasitol. 2009, 54, 180–185. [Google Scholar] [CrossRef]
- Miller, C.M.D.; Quinn, H.E.; Windsor, P.A.; Ellis, J.T. Characterisation of the first Australian isolate of Neospora caninum from cattle. Aust. Vet. J. 2002, 80, 620–625. [Google Scholar] [CrossRef]
- Okeoma, C.M.; Williamson, N.B.; Pomroy, W.E.; Stowell, K.M.; Gillespie, L.M. Isolation and molecular characterisation of Neospora caninum in cattle in New Zealand. NZ. Vet. J. 2004, 52, 364–370. [Google Scholar] [CrossRef]
- Fish, L.; Mazuz, M.; Molad, T.; Savitsky, I.; Shkap, V. Isolation of Neospora caninum from dairy zero grazing cattle in Israel. Vet. Parasitol. 2007, 149, 167–171. [Google Scholar] [CrossRef]
- Pastusiak, K.; Cabaj, W.; Moskwa, B. Isolation, identification and maintenance in cell culture of the first Polish isolate of Neospora caninum. Wiad. Parazytol. 2005, 51, 68. [Google Scholar]
- Goździk, K.; Cabaj, W. Characterization of the first Polish isolate of Neospora caninum from cattle. Acta Parasitol. 2007, 52, 295–297. [Google Scholar] [CrossRef]
- Rodrigues, A.A.R.; Gennari, S.M.; Aguiar, D.M.; Sreekumar, C.; Hill, D.E.; Miska, K.B.; Vianna, M.C.; Dubey, J.P. Shedding of Neospora caninum oocysts by dogs fed tissues from naturally infected water buffaloes (Bubalus bubalis) from Brazil. Vet. Parasitol. 2004, 124, 139–150. [Google Scholar] [CrossRef]
- Gondim, L.F.; McAllister, M.M.; Mateus-Pinilla, N.E.; Pitt, W.C.; Mech, L.D.; Nelson, M.E. Transmission of Neospora caninum between wild and domestic animals. J. Parasitol. 2004, 90, 1361–1365. [Google Scholar] [CrossRef]
- Vianna, M.C.; Sreekumar, C.; Miska, K.B.; Hill, D.E.; Dubey, J.P. Isolation of Neospora caninum from naturally infected white-tailed deer (Odocoileus virginianus). Vet. Parasitol. 2005, 129, 253–257. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P.; Duncan, R.B. Confirmation that the dog is a definitive host for Neospora caninum. Vet. Parasitol. 1999, 82, 327–333. [Google Scholar] [CrossRef]
- Gondim, L.F.; McAllister, M.M.; Pitt, W.C.; Zemlicka, D.E. Coyotes (Canis latrans) are definitive hosts of Neospora caninum. Int. J. Parasitol. 2004, 34, 159–161. [Google Scholar] [CrossRef]
- King, J.S.; Slapeta, J.; Jenkins, D.J.; Al-Qassab, S.E.; Ellis, J.T.; Windsor, P.A. Australian dingoes are definitive hosts of Neospora caninum. Int. J. Parasitol. 2010.
- Dubey, J.P.; Lindsay, D.S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 1996, 67, 1–59. [Google Scholar] [CrossRef]
- Dubey, J.P. Neosporosis: The first decade of research. Int. J. Parasitol. 1999, 29, 1485–1488. [Google Scholar] [CrossRef]
- Paré, J.; Thurmond, M.C.; Hietala, S.K. Congenital Neospora caninum infection in dairy cattle and associated calfhood mortality. Can. J. Vet. Res. 1996, 60, 133–139. [Google Scholar]
- Wouda, W.; Moen, A.R.; Schukken, Y.H. Abortion risk in progeny of cows after a Neospora caninum epidemic. Theriogenology 1998, 49, 1311–1316. [Google Scholar] [CrossRef]
- Dubey, J.P. Recent advances in Neospora and neosporosis. Vet. Parasitol. 1999, 84, 349–367. [Google Scholar] [CrossRef]
- Dijkstra, T.; Eysker, M.; Schares, G.; Conraths, F.J.; Wouda, W.; Barkema, H.W. Dogs shed Neospora caninum oocysts after ingestion of naturally infected bovine placenta but not after ingestion of colostrum spiked with Neospora caninum tachyzoites. Int. J. Parasitol. 2001, 31, 747–752. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Ritter, D.M.; Brake, D. Oocyst excretion in dogs fed mouse brains containing tissue cysts of a cloned line of Neospora caninum. J. Parasitol. 2001, 87, 909–911. [Google Scholar] [CrossRef]
- Dubey, J.P.; Liddell, S.; Mattson, D.; Speert, C.A.; Howe, D.K.; Jenkins, M.C. Characterization of the Oregon isolate of Neospora hughesi from a horse. J. Parasitol. 2001, 87, 345–353. [Google Scholar] [CrossRef]
- Dubey, J.P.; de Lahunta, A. Neosporosis associated congenital limb deformities in a calf. Appl. Parasitol. 1993, 34, 229–233. [Google Scholar]
- Bjerkås, I.; Jenkins, M.C.; Dubey, J.P. Identification and characterization of Neospora caninum tachyzoite antigens useful for diagnosis of Neosporosis. Clin. Diagn. Lab. Immunol. 1994, 1, 214–221. [Google Scholar]
- Lindsay, D.S.; Speer, C.A.; Toiviokinnucan, M.A.; Dubey, J.P.; Blagburn, B.L. Use of infected cultured cells to compare ultrastructural features of Neospora caninum from dogs and Toxoplasma gondii. Am. J. Vet. Res. 1993, 54, 103–106. [Google Scholar]
- Speer, C.A.; Dubey, J.P. Ultrastructure of tachyzoites, bradyzoites and tissue cysts of Neospora caninum. J. Raptor. Res. 1989, 36, 458–463. [Google Scholar]
- Speer, C.A.; Dubey, J.P.; McAllister, M.M.; Blixt, J.A. Comparative ultrastructure of tachyzoites, bradyzoites, and tissue cysts of Neospora caninum and Toxoplasma gondii. Int. J. Parasitol. 1999, 29, 1509–1519. [Google Scholar] [CrossRef]
- Barr, B.C.; Conrad, P.A.; Dubey, J.P.; Anderson, M.L. Neospora-like encephalomyelitis in a calf: pathology, ultrastructure, and immunoreactivity. J. Vet. Diagn. Invest. 1991, 3, 39–46. [Google Scholar] [CrossRef]
- McGuire, A.M.; McAllister, M.M.; Jolley, W.R.; Anderson-Sprecher, R.C. A protocol for the production of Neospora caninum tissue cysts in mice. J. Parasitol. 1997, 83, 647–651. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. Neospora caninum (Protozoa, Apicomplexa) infections in rats. Can. J. Zool. 1990, 68, 1595–1599. [Google Scholar] [CrossRef]
- Dubey, J.P.; Higgins, R.J.; Smith, J.H.; Otoole, T.D. Neospora caninum encephalomyelitis in a British dog. Vet. Record 1990, 126, 193–194. [Google Scholar]
- Barr, B.C.; Anderson, M.L.; Dubey, J.P.; Conrad, P.A. Neospora-like protozoal infections associated with bovine abortions. Vet. Pathol. 1991, 28, 110–116. [Google Scholar]
- Razmi, G.R.; Maleki, M.; Farzaneh, N.; Garoussi, M.T.; Fallah, A.H. First report of Neospora caninum-associated bovine abortion in Mashhad area, Iran. Parasitol. Res. 2007, 100, 755–757. [Google Scholar] [CrossRef]
- Barr, B.C.; Anderson, M.L.; Woods, L.W.; Dubey, J.P.; Conrad, P.A. Neospora-like protozoal infections associated with abortion in goats. J. Vet. Diagn. Invest. 1992, 4, 365–367. [Google Scholar] [CrossRef]
- Dubey, J.P.; Morales, J.A.; Villalobos, P.; Lindsay, D.S.; Blagburn, B.L.; Topper, M.J. Neosporosis-associated abortion in a dairy goat. J. Amer. Vet. Med. Assn. 1996, 208, 263–265. [Google Scholar]
- Basso, W.; Venturini, L.; Venturini, M.C.; Moore, P.; Rambeau, M.; Unzaga, J.M.; Campero, C.; Bacigalupe, D.; Dubey, J.R. Prevalence of Neospora caninum infection in dogs from beef-cattle farms, dairy farms, and from urban areas of Argentina. J. Parasitol. 2001, 87, 906–907. [Google Scholar] [CrossRef]
- McGuire, A.M.; McAllister, M.M.; Jolley, W.R. Separation and cryopreservation of Neospora caninum tissue cysts from murine brain. J. Parasitol. 1997, 83, 319–321. [Google Scholar]
- Lindsay, D.S.; Steinberg, H.; Dubielzig, R.R.; Semrad, S.D.; Konkle, D.M.; Miller, P.E.; Blagburn, B.L. Central nervous system neosporosis in a foal. J. Vet. Diagn. Invest. 1996, 8, 507–510. [Google Scholar] [CrossRef]
- Daft, B.M.; Barr, B.C.; Collins, N.; Sverlow, K. Neospora encephalomyelitis and polyradiculoneuritis in an aged mare with Cushing's disease. Equine Vet. J. 1997, 29, 240–243. [Google Scholar] [CrossRef]
- McCann, C.M.; McAllister, M.M.; Gondim, L.F.; Smith, R.F.; Cripps, P.J.; Kipar, A.; Williams, D.J.; Trees, A.J. Neospora caninum in cattle: experimental infection with oocysts can result in exogenous transplacental infection, but not endogenous transplacental infection in the subsequent pregnancy. Int. J. Parasitol. 2007, 37, 1631–1639. [Google Scholar] [CrossRef]
- Schares, G.; Heydorn, A.O.; Cüppers, A.; Conraths, F.J.; Mehlhorn, H. Cyclic transmission of Neospora caninum: serological findings in dogs shedding oocysts. Parasitol. Res. 2001, 87, 873–877. [Google Scholar] [CrossRef]
- McGarry, J.W.; Stockton, C.M.; Williams, D.J.; Trees, A.J. Protracted shedding of oocysts of Neospora caninum by a naturally infected foxhound. J. Parasitol. 2003, 89, 628–630. [Google Scholar] [CrossRef]
- Wapenaar, W.; Jenkins, M.C.; O'Handley, R.M.; Barkema, H.W. Neospora caninum-like oocysts observed in feces of free-ranging red foxes (Vulpes vulpes) and coyotes (Canis latrans). J. Parasitol. 2006, 92, 1270–1274. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. In vitro development of Neospora caninum (Protozoa: Apicomplexa) from dogs. J. Parasitol. 1989, 75, 163–165. [Google Scholar] [CrossRef]
- Thornton, R.N.; Thompson, E.J.; Dubey, J.P. Neospora abortion in New Zealand cattle. NZ. Vet. J. 1991, 39, 129–133. [Google Scholar] [CrossRef]
- Paré, J.; Hietala, S.K.; Thurmond, M.C. An enzyme-linked immunosorbent assay (ELISA) for serological diagnosis of Neospora sp. infection in cattle. J. Vet. Diagn. Invest. 1995, 7, 352–359. [Google Scholar] [CrossRef]
- Atkinson, R.; Harper, P.A.; Ryce, C.; Morrison, D.A.; Ellis, J.T. Comparison of the biological characteristics of two isolates of Neospora caninum. Parasitology 1999, 118, 363–370. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. Infections in mice with tachyzoites and bradyzoites of Neospora caninum (Protozoa: Apicomplexa). J. Parasitol. 1990, 76, 410–413. [Google Scholar] [CrossRef]
- Collantes-Fernández, E.; Arnaiz-Seco, I.; Burgos, B.M.; Rodriguez-Bertos, A.; Aduriz, G.; Fernandez-Garcia, A.; Ortega-Mora, L.M. Comparison of Neospora caninum distribution, parasite loads and lesions between epidemic and endemic bovine abortion cases. Vet. Parasitol. 2006, 142, 187–191. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Lenz, S.D.; Cole, R.A.; Dubey, J.P.; Blagburn, B.L. Mouse model for central nervous system Neospora caninum infections. J. Parasitol. 1995, 81, 313–315. [Google Scholar] [CrossRef]
- Quinn, H.E.; Miller, C.M.; Ryce, C.; Windsor, P.A.; Ellis, J.T. Characterization of an outbred pregnant mouse model of Neospora caninum infection. J. Parasitol. 2002, 88, 691–696. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lindsay, D.S. Gerbils (Meriones unguiculatus) are highly susceptible to oral infection with Neospora caninum oocysts. Parasitol. Res. 2000, 86, 165–168. [Google Scholar] [CrossRef]
- Williams, D.J.; Guy, C.S.; Smith, R.F.; Ellis, J.; Bjorkman, C.; Reichel, M.P.; Trees, A.J. Immunization of cattle with live tachyzoites of Neospora caninum confers protection against fetal death. Infec. Immunity 2007, 75, 1343–1348. [Google Scholar] [CrossRef]
- Rojo-Montejo, S.; Collantes-Fernandez, E.; Blanco-Murcia, J.; Rodriguez-Bertos, A.; Risco-Castillo, V.; Ortega-Mora, L.M. Experimental infection with a low virulence isolate of Neospora caninum at 70 days gestation in cattle did not result in foetopathy. Vet. Res. 2009, 40, 49. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lindsay, D.S. Neospora caninum induced abortion in sheep. J. Vet. Diagn. Invest. 1990, 2, 230–233. [Google Scholar] [CrossRef]
- McAllister, M.M.; McGuire, A.M.; Jolley, W.R.; Lindsay, D.S.; Trees, A.J.; Stobart, R.H. Experimental neosporosis in pregnant ewes and their offspring. Vet. Pathol. 1996, 33, 647–655. [Google Scholar]
- Buxton, D.; Maley, S.W.; Thomson, K.M.; Trees, A.J.; Innes, E.A. Experimental infection of non-pregnant and pregnant sheep with Neospora caninum. J. Comp. Pathol. 1997, 117, 1–16. [Google Scholar]
- Buxton, D.; Maley, S.W.; Wright, S.; Thomson, K.M.; Rae, A.G.; Innes, E.A. The pathogenesis of experimental neosporosis in pregnant sheep. J. Comp. Pathol. 1998, 118, 267–279. [Google Scholar] [CrossRef]
- Buxton, D.; Wright, S.; Maley, S.W.; Rae, A.G.; Lundén, A.; Innes, E.A. Immunity to experimental neosporosis in pregnant sheep. Parasite Immunol. 2001, 23, 85–91. [Google Scholar]
- Howe, D.K.; Crawford, A.C.; Lindsay, D.; Sibley, L.D. The p29 and p35 immunodominant antigens of Neospora caninum tachyzoites are homologous to the family of surface antigens of Toxoplasma gondii. Infec. Immunity 1998, 66, 5322–5328. [Google Scholar]
- Marsh, A.E.; Howe, D.K.; Wang, G.; Barr, B.C.; Cannon, N.; Conrad, P.A. Differentiation of Neospora hughesi from Neospora caninum based on their immunodominant surface antigen, SAG1 and SRS2. Int. J. Parasitol. 1999, 29, 1575–1582. [Google Scholar] [CrossRef]
- Shin, Y.S.; Lee, E.G.; Jung, T.S. Exploration of immunoblot profiles of Neospora caninum probed with different bovine immunoglobulin classes. J. Vet. Sci. 2005, 6, 157–160. [Google Scholar]
- Lee, E.G.; Kim, J.H.; Shin, Y.S.; Shin, G.W.; Kim, Y.R.; Palaksha, K.J.; Kim, D.Y.; Yamane, I.; Kim, Y.H.; Kim, G.S.; Suh, M.D.; Jung, T.S. Application of proteomics for comparison of proteome of Neospora caninum and Toxoplasma gondii tachyzoites. J. Chromat. B 2005, 815, 305–314. [Google Scholar] [CrossRef]
- Morrison, D.A.; Ellis, J.T. Effects of nucleotide sequence alignment on phylogeny estimation: a case study of 18S rDNAs of apicomplexa. Mol Biol Evol 1997, 14, 428–441. [Google Scholar] [CrossRef]
- Marsh, A.E.; Barr, B.C.; Sverlow, K.; Ho, M.; Dubey, J.P.; Conrad, P.A. Sequence analysis and comparison of ribosomal DNA from bovine Neospora to similar coccidial parasites. J. Parasitol. 1995, 81, 530–535. [Google Scholar] [CrossRef]
- Holmdahl, J.; Björkman, C.; Stenlund, S.; Uggla, A.; Dubey, J.P. Bovine Neospora and Neospora caninum: One and the same. Parasitol. Today 1997, 13, 40–41. [Google Scholar]
- Johnson, A.M.; Illana, S.; Dubey, J.P.; Dame, J.B. Toxoplasma gondii and Hammondia hammondi: DNA comparison using cloned rRNA gene probes. Exp. Parasitol. 1987, 63, 272–278. [Google Scholar] [CrossRef]
- Gondim, L.F.; Laski, P.; Gao, L.; McAllister, M.M. Variation of the internal transcribed spacer 1 sequence within individual strains and among different strains of Neospora caninum. J. Parasitol. 2004, 90, 119–122. [Google Scholar] [CrossRef]
- Spencer, J.A.; Witherow, A.K.; Blagburn, B.L. A random amplified polymorphic DNA polymerase chain reaction technique that differentiates between Neospora species. J. Parasitol. 2000, 86, 1366–1368. [Google Scholar] [CrossRef]
- Regidor-Cerrillo, J.; Pedraza-Díaz, S.; Gómez-Bautista, M.; Ortega-Mora, L.M. Multilocus microsatellite analysis reveals extensive genetic diversity in Neospora caninum. J. Parasitol. 2006, 92, 517–524. [Google Scholar] [CrossRef]
- Kaufmann, H.; Yamage, M.; Roditi, I.; Dobbelaere, D.; Dubey, J.P.; Holmdahl, O.J.; Trees, A.; Gottstein, B. Discrimination of Neospora caninum from Toxoplasma gondii and other apicomplexan parasites by hybridization and PCR. Mol. Cell. Probe. 1996, 10, 289–297. [Google Scholar]
- Pedraza-Diaz, S.; Marugan-Hernandez, V.; Collantes-Fernandez, E.; Regidor-Cerrillo, J.; Rojo-Montejo, S.; Gomez-Bautista, M.; Ortega-Mora, L.M. Microsatellite markers for the molecular characterization of Neospora caninum: application to clinical samples. Vet. Parasitol. 2009, 166, 38–46. [Google Scholar] [CrossRef]
- Al-Qassab, S.; Reichel, M.P.; Ivens, A.; Ellis, J.T. Genetic diversity amongst isolates of Neospora caninum, and the development of a multiplex assay for the detection of distinct strains. Mol. Cell. Probe. 2009, 23, 132–139. [Google Scholar] [CrossRef]
- Al-Qassab, S.; Reichel, M.P.; Ellis, J. A second generation multiplex PCR for typing strains of Neospora caninum using six DNA targets. Mol. Cell. Probe. 2009, 24, 20–26. [Google Scholar] [CrossRef]
- Siverajah, S.; Ryce, C.; Morrison, D.A.; Ellis, J.T. Characterization of an alpha tubulin gene sequence from Neospora caninum and Hammondia heydorni, and their comparison to homologous genes from Apicomplexa. Parasitology 2003, 126, 561–569. [Google Scholar]
- Ellis, J.T.; Ryce, C.; Atkinson, R.; Balu, S.; Holmdahl, O.J.M. Neospora caninum: gene discovery through analysis of expressed sequence taqs. Int. J. Parasitol. 2000, 30, 909–913. [Google Scholar]
- Fernández-García, A.; Risco-Castillo, V.; Zaballos, A.; Alvarez-García, G.; Ortega-Mora, L.M. Identification and molecular cloning of the Neospora caninum SAG4 gene specifically expressed at bradyzoite stage. Mol. Biochem. Parasitol. 2006, 146, 89–97. [Google Scholar] [CrossRef]
- Walsh, C.P.; Vemulapalli, R.; Sriranganathan, N.; Zajac, A.M.; Jenkins, M.C.; Lindsay, D.S. Molecular comparison of the dense granule proteins GRA6 and GRA7 of Neospora hughesi and Neospora caninum. Int. J. Parasitol. 2001, 31, 253–258. [Google Scholar] [CrossRef]
- Innes, E.A.; Panton, W.R.M.; Marks, J.; Trees, A.J.; Holmdahl, J.; Buxton, D. Interferon-gamma inhibits the intracellular multiplication of Neospora caninum, as shown by incorporation of ³[H] uracil. J. Comp. Pathol. 1995, 113, 95–100. [Google Scholar] [CrossRef]
- Pérez-Zaballos, F.J.; Ortega-Mora, L.M.; Álvarez-García, G.; Collantes-Fernández, E.; Navarro-Lozano, V.; García-Villada, L.; Costas, E. Adaptation of Neospora caninum isolates to cell-culture changes: An argument in favor of its clonal population structure. J. Parasitol. 2005, 91, 507–510. [Google Scholar] [CrossRef]
- Dubey, J.P.; Barr, B.C.; Barta, J.R.; Bjerkås, I.; Björkman, C.; Blagburn, B.L.; Bowman, D.D.; Buxton, D.; Ellis, J.T.; Gottstein, B.; Hemphill, A.; Hill, D.E.; Howe, D.K.; Jenkins, M.C.; Kobayashi, Y.; Koudela, B.; Marsh, A.E.; Mattsson, J.G.; McAllister, M.M.; Modrý, D.; Omata, Y.; Sibley, L.D.; Speer, C.A.; Trees, A.J.; Uggla, A.; Upton, S.J.; Williams, D.J.L.; Lindsay, D.S. Redescription of Neospora caninum and its differentiation from related coccidia. Int. J. Parasitol. 2002, 32, 929–946. [Google Scholar] [CrossRef]
- Collantes-Fernández, E.; Lopez-Perez, I.; Álvarez-García, G.; Ortega-Mora, L.M. Temporal distribution and parasite load kinetics in blood and tissues during Neospora caninum infection in mice. Infec. Immunity 2006, 74, 2491–2494. [Google Scholar] [CrossRef]
- Shin, Y.S.; Shin, G.W.; Kim, Y.R.; Lee, E.Y.; Yang, H.H.; Palaksha, K.J.; Youn, H.J.; Kim, J.H.; Kim, D.Y.; Marsh, A.E.; Lakritz, J.; Jung, T.S. Comparison of proteome and antigenic proteome between two Neospora caninum isolates. Vet. Parasitol. 2005, 134, 41–52. [Google Scholar] [CrossRef]
- Lehmann, T.; Marcet, P.L.; Graham, D.H.; Dahl, E.R.; Dubey, J.P. Globalization and the population structure of Toxoplasma gondii. Proc.Nat.Acad.Sci.USA 2006, 103, 11423–11428. [Google Scholar]
- Müller, N.; Sager, H.; Hemphill, A.; Mehlhorn, H.; Heydorn, A.O.; Gottstein, B. Comparative molecular investigation of Nc5-PCR amplicons from Neospora caninum NC-1 and Hammondia heydorni-Berlin-1996. Parasitol. Res. 2001, 87, 883–885. [Google Scholar] [CrossRef]
- Yamage, M.; Flechtner, O.; Gottstein, B. Neospora caninum: specific oligonucleotide primers for the detection of brain "cyst" DNA of experimentally infected nude mice by the polymerase chain reaction (PCR). J. Parasitol. 1996, 82, 272–279. [Google Scholar] [CrossRef]
- Gasser, R.B. Molecular tools--advances, opportunities and prospects. Vet. Parasitol. 2006, 136, 69–89. [Google Scholar] [CrossRef]
- Shibahara, T.; Kokuho, T.; Eto, M.; Haritani, M.; Hamaoka, T.; Shimura, K.; Nakamura, K.; Yokomizo, Y.; Yamane, I. Pathological and immunological findings of athymic nude and congenic wild type BALB/c mice experimentally infected with Neospora caninum. Vet. Pathol. 1999, 36, 321–327. [Google Scholar]
- Reichel, M.P.; Ellis, J.T. Neospora caninum—how close are we to development of an efficacious vaccine that prevents abortion in cattle? Int. J. Parasitol. 2009, 39, 1173–1187. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Al-Qassab, S.E.; Reichel, M.P.; Ellis, J.T. On the Biological and Genetic Diversity in Neospora caninum . Diversity 2010, 2, 411-438. https://doi.org/10.3390/d2030411
Al-Qassab SE, Reichel MP, Ellis JT. On the Biological and Genetic Diversity in Neospora caninum . Diversity. 2010; 2(3):411-438. https://doi.org/10.3390/d2030411
Chicago/Turabian StyleAl-Qassab, Sarwat E., Michael P. Reichel, and John T. Ellis. 2010. "On the Biological and Genetic Diversity in Neospora caninum " Diversity 2, no. 3: 411-438. https://doi.org/10.3390/d2030411