The Interactive Effect of Tonic Pain and Motor Learning on Corticospinal Excitability


Abstract
:1. Introduction
2. Methods
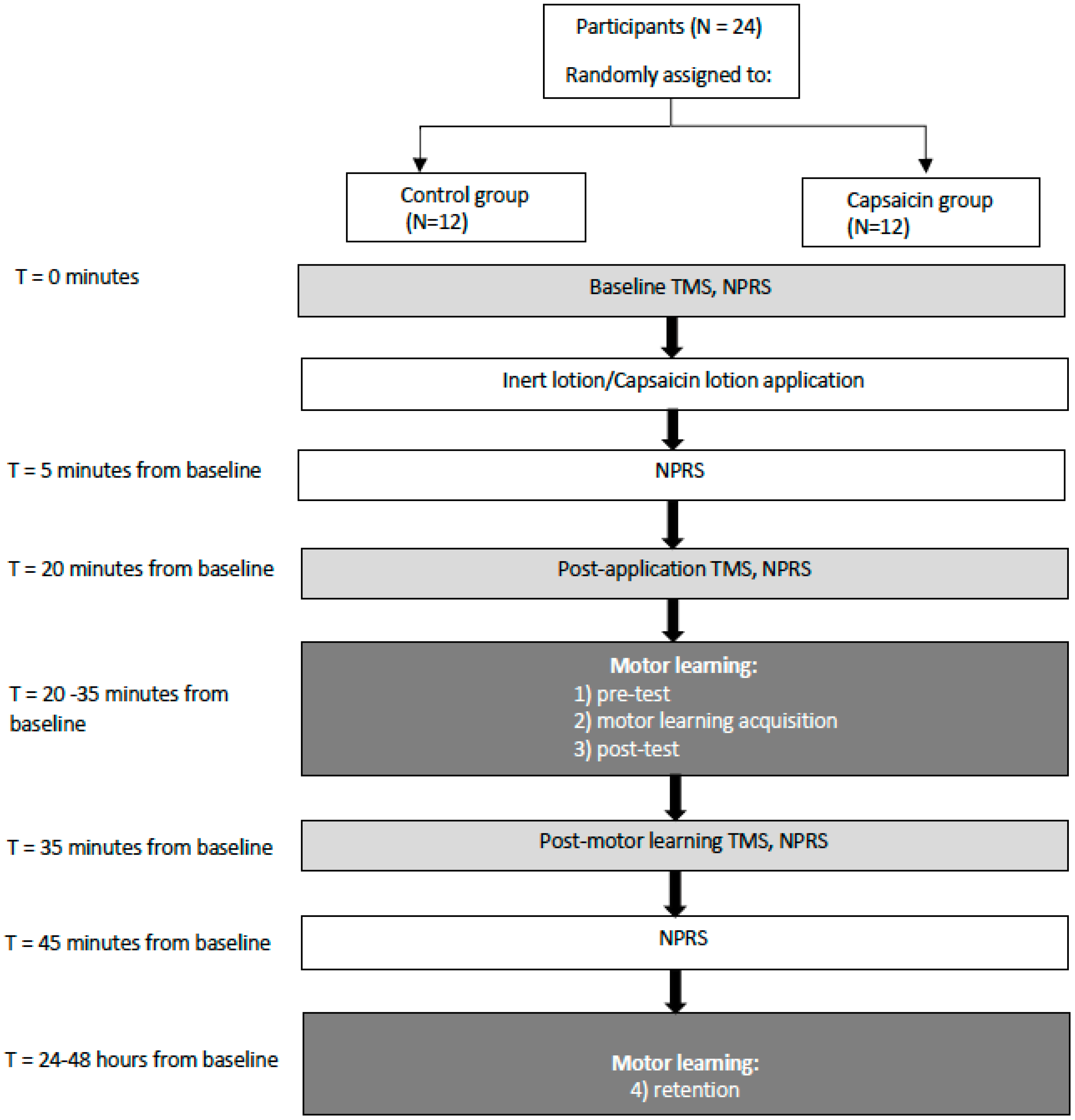
2.1. Methods Overview
2.2. Outcome Measures
2.2.1. Pain
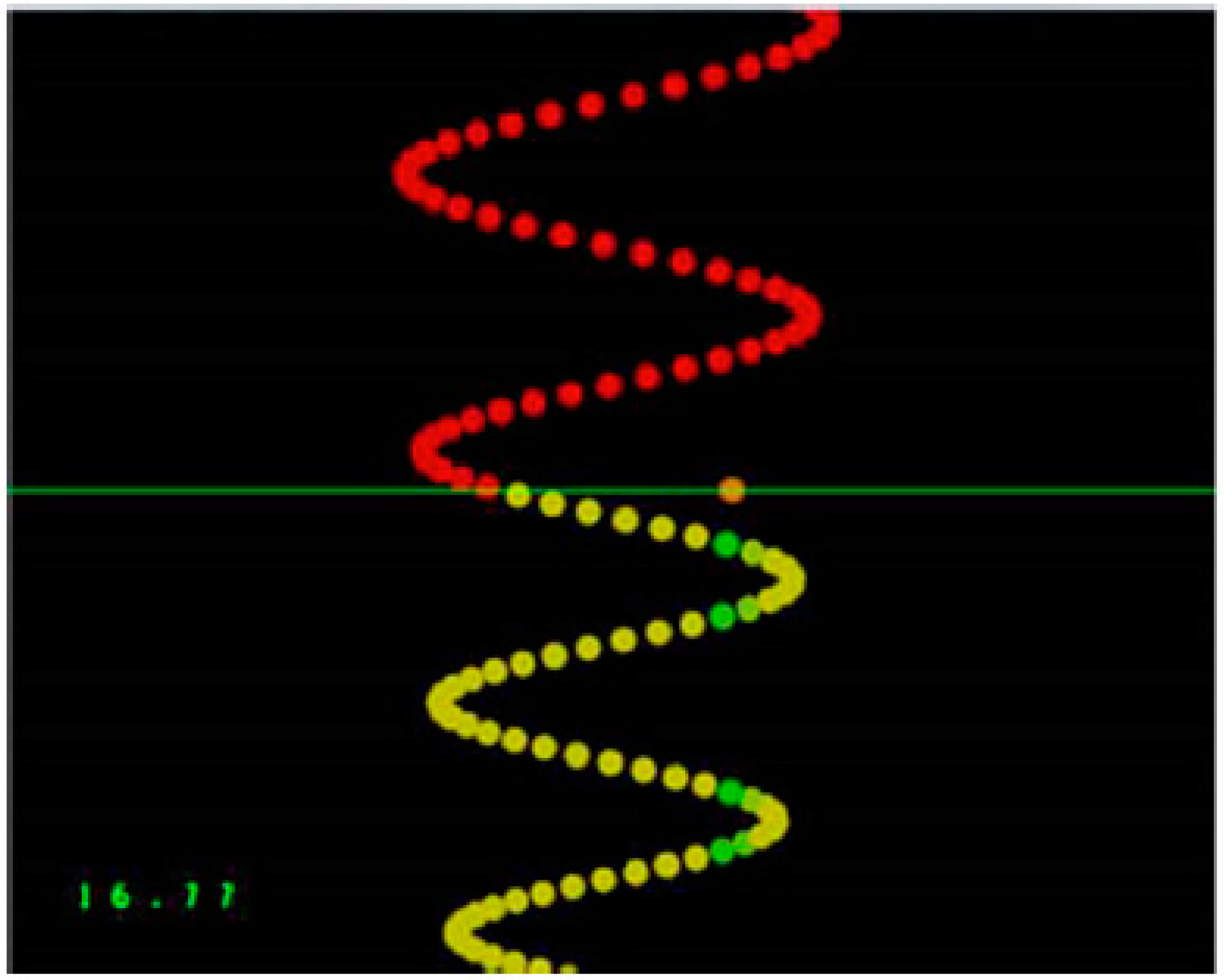
2.2.2. Motor Learning Task
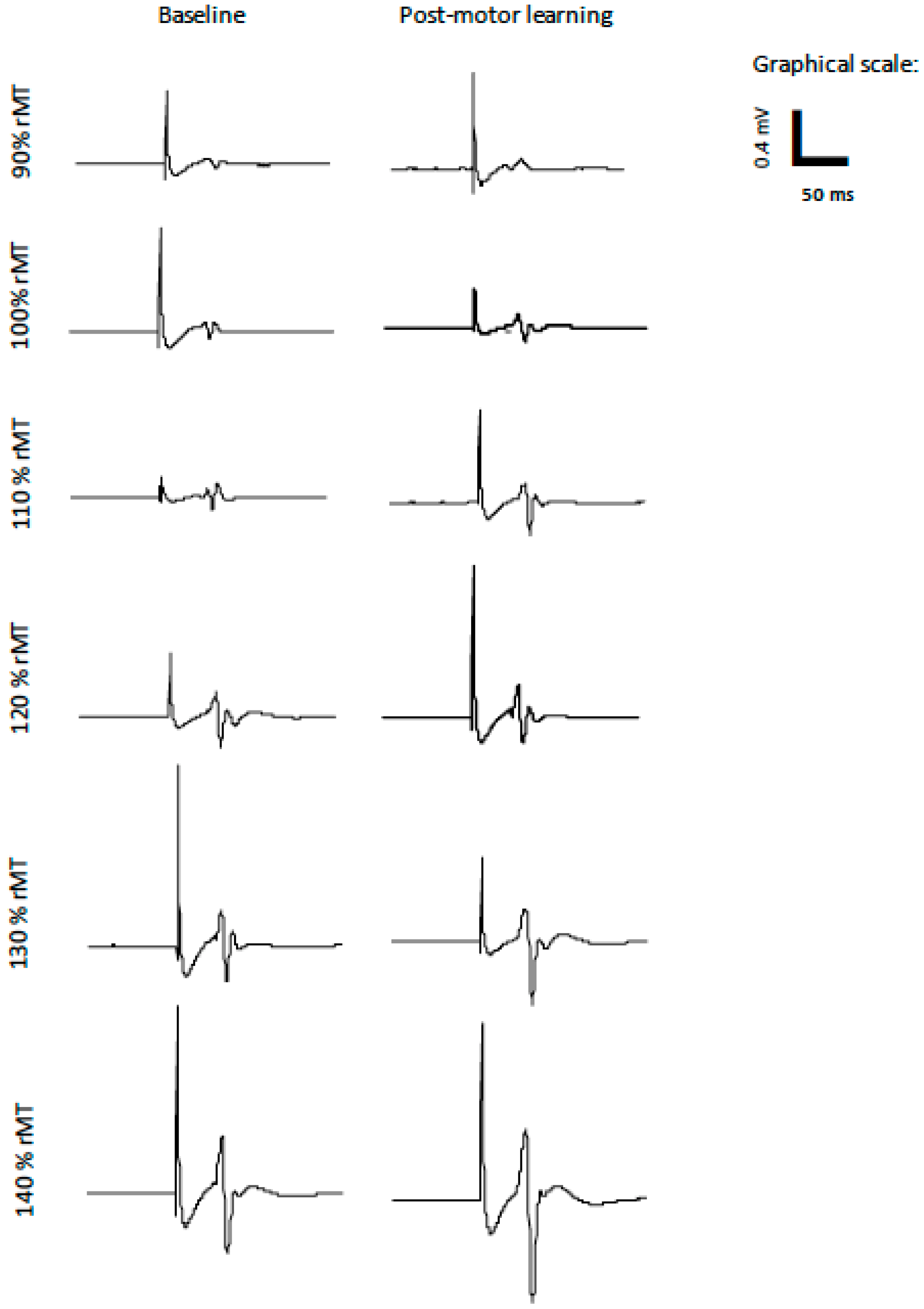
2.2.3. Transcranial Magnetic Stimulation
2.2.4. Electromyography Recording
2.2.5. IO Curves
2.3. Data Analysis
3. Results
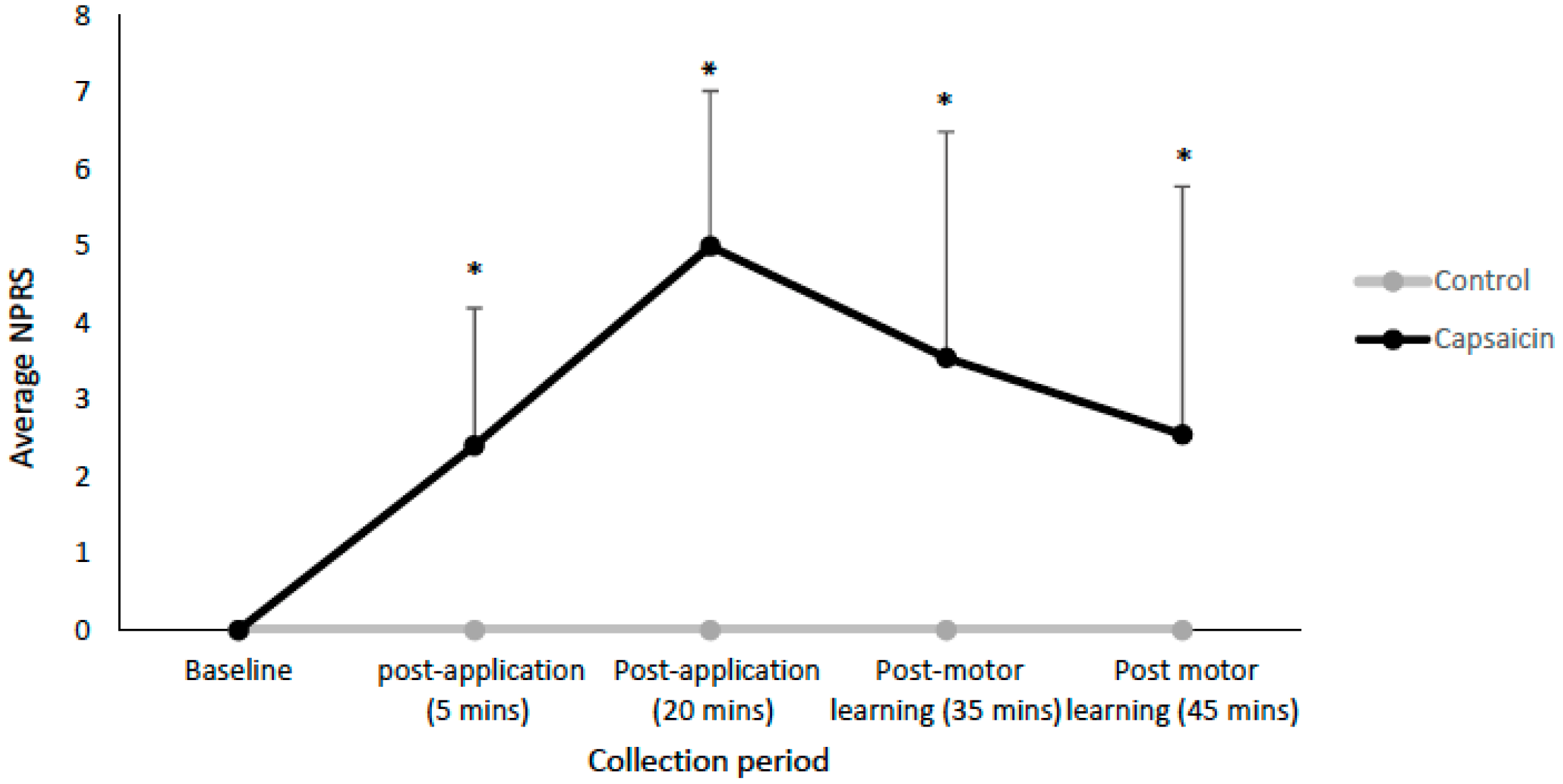
3.1. Pain Ratings
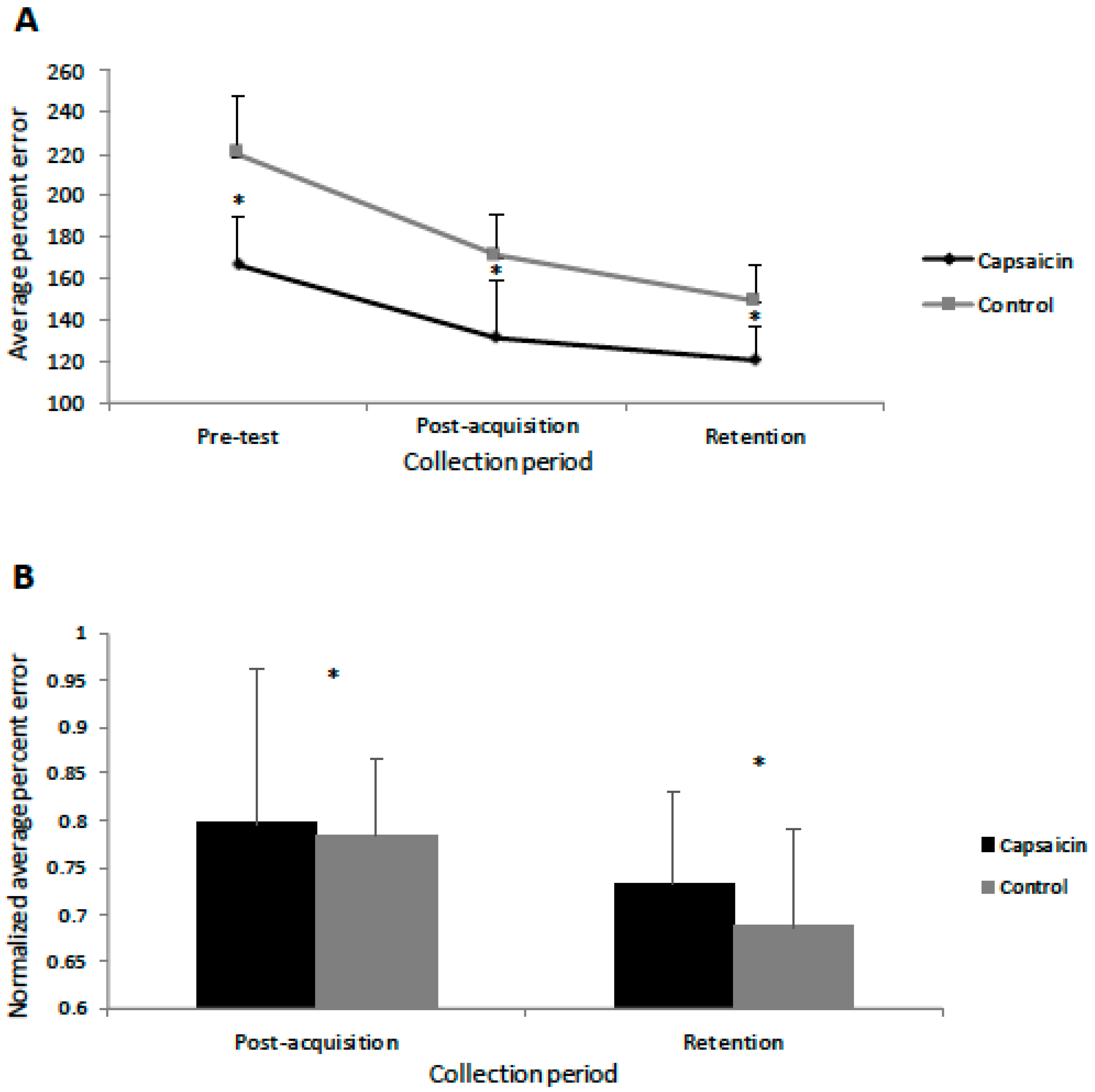
3.2. Behavioural Data
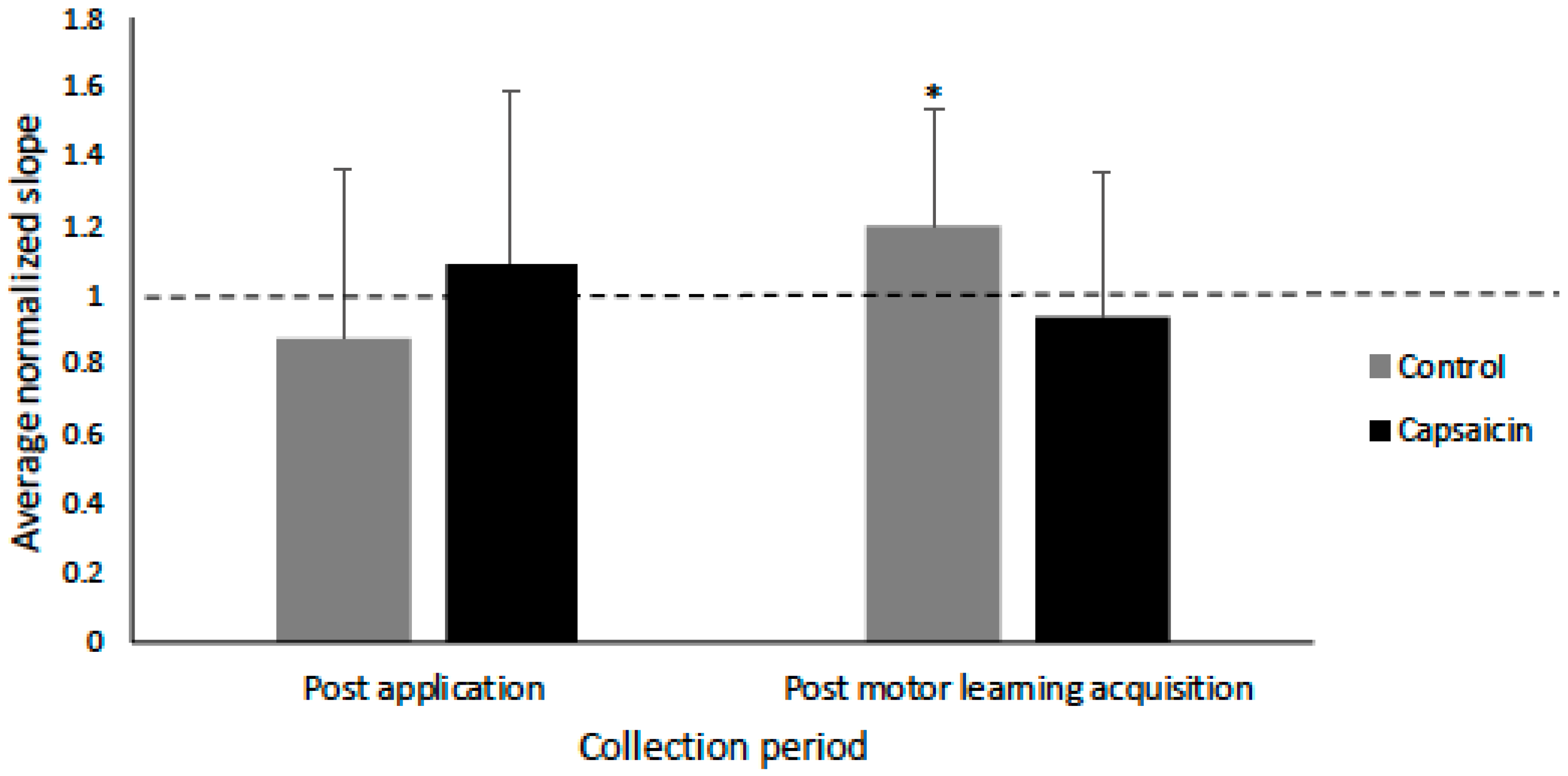
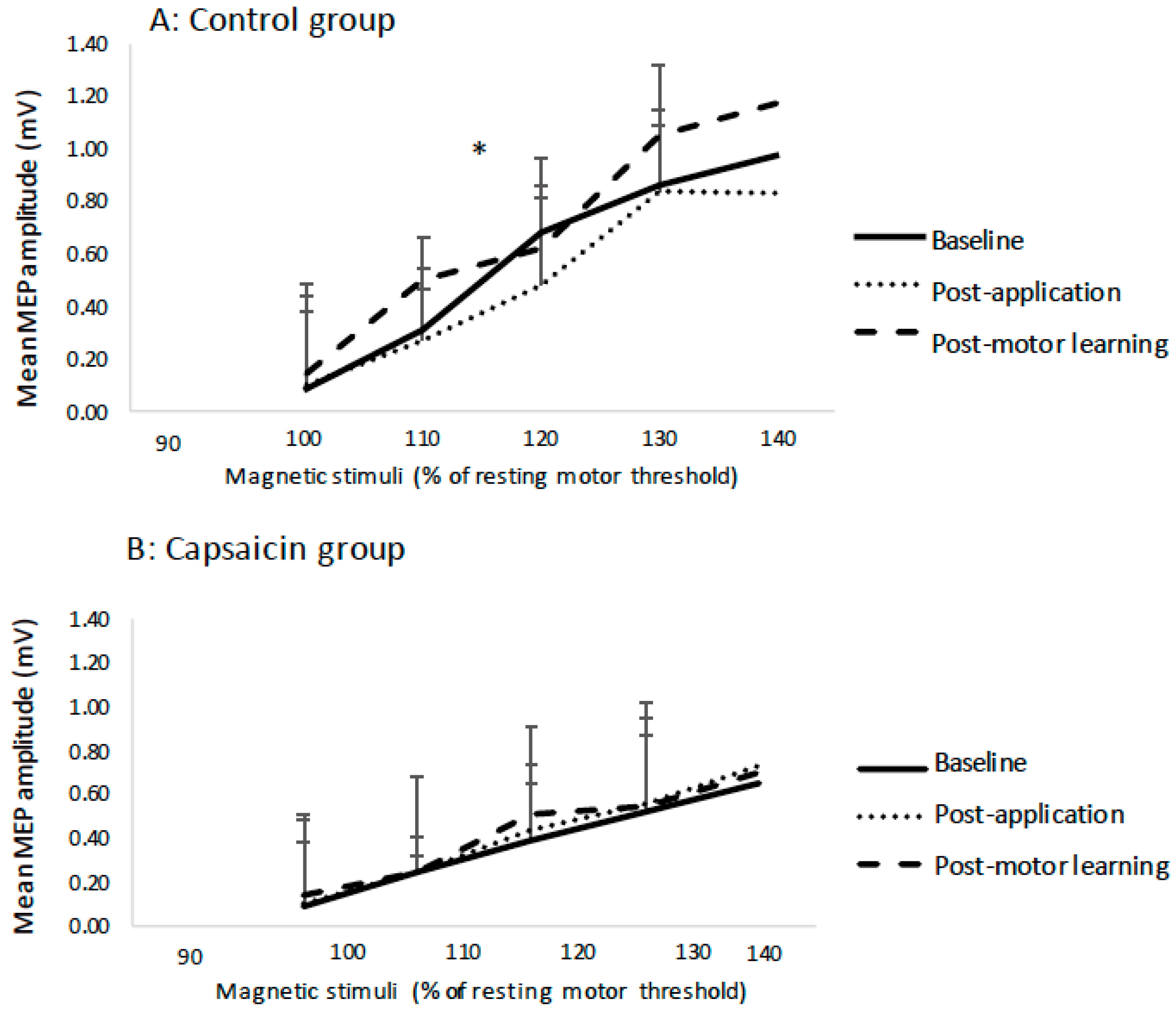
3.3. Transcranial IO Curves
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hodges, P.W.; Tucker, K. Moving differently in pain: A new theory to explain the adaptation to pain. Pain 2011, 152, S90–S98. [Google Scholar] [CrossRef]
- Bank, P.; Peper, C.; Marinus, J.; Beek, P.; Hilten, J. Motor consequences of experimentally induced limb pain: A systematic review. Eur. J. Pain 2013, 17, 145–157. [Google Scholar] [CrossRef]
- Dancey, E.; Murphy, B.; Srbely, J.; Yielder, P. The effect of experimental pain on motor training performance and sensorimotor integration. Exp. Brain Res. 2014, 232, 2879–2889. [Google Scholar] [CrossRef] [Green Version]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The effect of local versus remote experimental pain on motor learning and sensorimotor integration using a complex typing task. Pain 2016, 157, 1682–1695. [Google Scholar] [CrossRef]
- Sanes, J.N.; Donoghue, J.P. Plasticity and primary motor cortex. Annu. Rev. Neurosci. 2000, 23, 393–415. [Google Scholar] [CrossRef]
- Karni, A.; Meyer, G.; Jezzard, P.; Adams, M.M.; Turner, R.; Ungerleider, L.G. Functional MRI evidence For adult motor cortex plasticity during motor skill learning. Nature 1995, 377, l55–l58. [Google Scholar]
- Nudo, R.; Milliken, G.; Jenkins, W.M.; Merzenich, M. Use-dependent alterations of movement representations in primary motor cortex of adult squirrel monkeys. J. Neurosci. 1996, 16, 785–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Leone, A.; Nguyet, D.; Cohen, L.G.; Brasil-Neto, J.P.; Cammarota, A.; Hallett, M. Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition of new fine motor skills. J. Neurophysiol. 1995, 74, 1037–1045. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Grafman, J.; Hallett, M. Modulation of cortical motor output maps during development of implicit and explicit knowledge. Science 1994, 263, 1287–1289. [Google Scholar] [CrossRef]
- Classen, J.; Liepert, J.; Wise, S.; Hallett, M.; Cohen, L. Rapid plasticity of human cortical movement representation induced by practice. J. Neurophysiol. 1998, 79, 1117–1123. [Google Scholar] [CrossRef]
- Bütefisch, C.M.; Davis, B.C.; Wise, S.P.; Sawaki, L.; Kopylev, L.; Classen, J.; Cohen, L.G. Mechanisms of use-dependent plasticity in the human motor cortex. Proc. Natl. Acad. Sci. USA 2000, 97, 3661–3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flor, H. Cortical reorganisation and chronic pain: Implications for rehabilitation. J. Rehabil. Med.-Suppl. 2003, 35, 66–72. [Google Scholar] [CrossRef]
- Schweinhardt, P.; Lee, M.; Tracey, I. Imaging pain in patients: Is it meaningful? Curr. Opin. Neurol. 2006, 19, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, S.; Romaniello, A.; Wang, K.; Svensson, P.; Sessle, B.J.; Arendt-Nielsen, L. The effects of intra-oral pain on motor cortex neuroplasticity associated with short-term novel tongue-protrusion training in humans. Pain 2007, 132, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, M.-C.; Roosink, M.; Mercier, C. Effect of local versus remote tonic heat pain during training on acquisition and retention of a finger-tapping sequence task. Exp. Brain Res. 2015, 234, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Bouffard, J.; Bouyer, L.J.; Roy, J.-S.; Mercier, C. Tonic pain experienced during locomotor training impairs retention despite normal performance during acquisition. J. Neurosci. 2014, 34, 9190–9195. [Google Scholar] [CrossRef]
- Rittig-Rasmussen, B.; Kasch, H.; Fuglsang-Frederiksen, A.; Svensson, P.; Jensen, T.S. The role of neuroplasticity in experimental neck pain: A study of potential mechanisms impeding clinical outcomes of training. Man. Ther. 2014, 19, 288–293. [Google Scholar] [CrossRef]
- McGaughy, J.; Dalley, J.; Morrison, C.; Everitt, B.; Robbins, T. Selective behavioral and neurochemical effects of cholinergic lesions produced by intrabasalis infusions of 192 IgG-saporin on attentional performance in a five-choice serial reaction time task. J. Neurosci. 2002, 22, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Culberson, A.; Packowski, C.; Chiba, A.A.; Tuszynski, M.H. Lesions of the basal forebrain cholinergic system impair task acquisition and abolish cortical plasticity associated with motor skill learning. Neuron 2003, 38, 819–829. [Google Scholar] [CrossRef]
- Rosenkranz, K.; Rothwell, J.C. The effect of sensory input and attention on the sensorimotor organization of the hand area of the human motor cortex. J. Physiol. 2004, 561, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Stefan, K.; Wycislo, M.; Classen, J. Modulation of associative human motor cortical plasticity by attention. J. Neurophysiol. 2004, 92, 66–72. [Google Scholar] [CrossRef]
- Coull, J.T. Neural correlates of attention and arousal: Insights from electrophysiology, functional neuroimaging and psychopharmacology. Prog. Neurobiol. 1998, 55, 343–361. [Google Scholar] [CrossRef]
- Treede, R.-D.; Magerl, W. Multiple mechanisms of secondary hyperalgesia. Prog. Brain Res. 2000, 129, 331–341. [Google Scholar]
- Koltzenburg, M.; Torebjörk, H.E.; Wahren, L.K. Nociceptor modulated central sensitization causes mechanical hyperalgesia in acute chemogenic and chronic neuropathic pain. Brain 1994, 117, 579–591. [Google Scholar] [CrossRef]
- Maihöfner, C.; Handwerker, H.O. Differential coding of hyperalgesia in the human brain: A functional MRI study. Neuroimage 2005, 28, 996–1006. [Google Scholar] [CrossRef]
- Torebjörk, H.; Lundberg, L.; LaMotte, R. Central changes in processing of mechanoreceptive input in capsaicin-induced secondary hyperalgesia in humans. J. Physiol. 1992, 448, 765–780. [Google Scholar] [CrossRef]
- Schmidt, R.; Schmelz, M.; Forster, C.; Ringkamp, M.; Torebjork, E.; Handwerker, H. Novel classes of responsive and unresponsive C nociceptors in human skin. J. Neurosci. 1995, 15, 333–341. [Google Scholar] [CrossRef]
- Dettmers, C.; Adler, T.; Rzanny, R.; Van Schayck, R.; Gaser, C.; Weiss, T.; Miltner, W.; Brückner, L.; Weiller, C. Increased excitability in the primary motor cortex and supplementary motor area in patients with phantom limb pain after upper limb amputation. Neurosci. Lett. 2001, 307, 109–112. [Google Scholar] [CrossRef]
- Krause, P.; Förderreuther, S.; Straube, A. TMS motor cortical brain mapping in patients with complex regional pain syndrome type I. Clin. Neurophysiol. 2006, 117, 169–176. [Google Scholar] [CrossRef]
- Falla, D.; Farina, D. Neuromuscular adaptation in experimental and clinical neck pain. J. Electromyogr. Kinesiol. 2008, 18, 255–261. [Google Scholar] [CrossRef]
- Madeleine, P.; Leclerc, F.; Arendt-Nielsen, L.; Ravier, P.; Farina, D. Experimental muscle pain changes the spatial distribution of upper trapezius muscle activity during sustained contraction. Clin. Neurophysiol. 2006, 117, 2436–2445. [Google Scholar] [CrossRef]
- Madeleine, P.; Lundager, B.; Voigt, M.; Arendt-Nielsen, L. Shoulder muscle co-ordination during chronic and acute experimental neck-shoulder pain. An occupational pain study. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 79, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Sae-Lee, D.; Whittle, T.; Forte, A.R.; Peck, C.C.; Byth, K.; Sessle, B.J.; Murray, G.M. Effects of experimental pain on jaw muscle activity during goal-directed jaw movements in humans. Exp. Brain Res. 2008, 189, 451–462. [Google Scholar] [CrossRef]
- Romaniello, A.; Cruccu, G.; McMillan, A.S.; Arendt-Nielsen, L.; Svensson, P. Effect of experimental pain from trigeminal muscle and skin on motor cortex excitability in humans. Brain Res. 2000, 882, 120–127. [Google Scholar] [CrossRef]
- Valeriani, M.; Restuccia, D.; Di Lazzaro, V.; Oliviero, A.; Profice, P.; Le Pera, D.; Saturno, E.; Tonali, P. Inhibition of the human primary motor area by painful heat stimulation of the skin. Clin. Neurophysiol. 1999, 110, 1475–1480. [Google Scholar] [CrossRef]
- Valeriani, M.; Restuccia, D.; Di Lazzaro, V.; Oliviero, A.; Le Pera, D.; Profice, P.; Saturno, E.; Tonali, P. Inhibition of biceps brachii muscle motor area by painful heat stimulation of the skin. Exp. Brain Res. 2001, 139, 168–172. [Google Scholar] [CrossRef]
- Le Pera, D.; Graven-Nielsen, T.; Valeriani, M.; Oliviero, A.; Di Lazzaro, V.; Tonali, P.A.; Arendt-Nielsen, L. Inhibition of motor system excitability at cortical and spinal level by tonic muscle pain. Clin. Neurophysiol. 2001, 112, 1633–1641. [Google Scholar] [CrossRef]
- Hirano, M.; Kubota, S.; Tanabe, S.; Koizume, Y.; Funase, K. Interactions among learning stage, retention, and primary motor cortex excitability in motor skill learning. Brain Stimul. 2015, 8, 1195–1204. [Google Scholar] [CrossRef]
- Muellbacher, W.; Ziemann, U.; Boroojerdi, B.; Cohen, L.; Hallett, M. Role of the human motor cortex in rapid motor learning. Exp. Brain Res. 2001, 136, 431–438. [Google Scholar] [CrossRef]
- Karni, A. The acquisition of perceptual and motor skills: A memory system in the adult human cortex. Cogn. Brain Res. 1996, 5, 39–48. [Google Scholar] [CrossRef]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The interactive effect of acute pain and motor learning acquisition on sensorimotor integration and motor learning outcomes. J. Neurophysiol. 2016, 116, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Yielder, P.; Murphy, B. Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration? Brain Sci. 2018, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Ingham, D.; Tucker, K.J.; Tsao, H.; Hodges, P.W. The effect of pain on training-induced plasticity of the corticomotor system. Eur. J. Pain 2011, 15, 1028–1034. [Google Scholar] [CrossRef]
- Mavromatis, N.; Neige, C.; Gagné, M.; Reilly, K.T.; Mercier, C. Effect of Experimental Hand Pain on Training-Induced Changes in Motor Performance and Corticospinal Excitability. Brain Sci. 2017, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.; Lavender, A.P.; Ridding, M.C.; Semmler, J.G. Motor cortex plasticity induced by paired associative stimulation is enhanced in physically active individuals. J. Physiol. 2009, 587, 5831–5842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebner, H.; Rothwell, J. Transcranial magnetic stimulation: New insights into representational cortical plasticity. Exp. Brain Res. 2003, 148, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dolphin, N.W.; Crue, B.L., Jr. Pain: Clinical Manual For Nursing Practice. Clin. J. Pain 1989, 5, 363. [Google Scholar] [CrossRef]
- Holland, L.; Murphy, B.; Passmore, S.; Yielder, P. Time course of corticospinal excitability changes following a novel motor training task. Neurosci. Lett. 2015, 591, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Kawai, S.; Fuchigami, Y.; Morita, H.; Ofuji, A. The effect of current direction induced by transcranial magnetic stimulation on the corticospinal excitability in human brain. Electroencephalogr. Clin. Neurophysiol./Electromyogr. Mot. Control 1996, 101, 478–482. [Google Scholar]
- Werhahn, K.; Fong, J.; Meyer, B.-U.; Priori, A.; Rothwell, J.; Day, B.; Thompson, P. The effect of magnetic coil orientation on the latency of surface EMG and single motor unit responses in the first dorsal interosseous muscle. Electroencephalogr. Clin. Neurophysiol./Evoked Potentials Sect. 1994, 93, 138–146. [Google Scholar] [CrossRef]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.; George, M. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an IFCN Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Pennisi, G.; Bella, R.; Lanza, G. Motor cortex plasticity in subcortical ischemic vascular dementia: What can TMS say? Clin. Neurophysiol. 2015, 126, 851–852. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.; Rogasch, N.C.; Semmler, J.G. Hemispheric differences in use-dependent corticomotor plasticity in young and old adults. Exp. Brain Res. 2010, 205, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Muellbacher, W.; Hallett, M.; Cohen, L.G. Modulation of practice-dependent plasticity in human motor cortex. Brain 2001, 124, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muellbacher, W.; Ziemann, U.; Wissel, J.; Dang, N.; Kofler, M.; Facchini, S.; Boroojerdi, B.; Poewe, W.; Hallett, M. Early consolidation in human primary motor cortex. Nature 2002, 415, 640–644. [Google Scholar] [CrossRef]
- Rossi, F.; Triggs, W.J.; Eisenschenk, S. Topographic differences in task-dependent facilitation of magnetic motor evoked potentials. Neurology 1999, 52, 537. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, K.; Kacar, A.; Rothwell, J.C. Differential modulation of motor cortical plasticity and excitability in early and late phases of human motor learning. J. Neurosci. 2007, 27, 12058–12066. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J. Techniques and mechanisms of action of transcranial stimulation of the human motor cortex. J. Neurosci. Methods 1997, 74, 113–122. [Google Scholar] [CrossRef]
- Wolpaw, J.R.; Tennissen, A.M. Activity-dependent spinal cord plasticity in health and disease. Annu. Rev. Neurosci. 2001, 24, 807–843. [Google Scholar] [CrossRef]
- Thompson, A.K.; Chen, X.Y.; Wolpaw, J.R. Acquisition of a simple motor skill: Task-dependent adaptation plus long-term change in the human soleus H-reflex. J. Neurosci. 2009, 29, 5784–5792. [Google Scholar] [CrossRef]
- Hess, G.; Donoghue, J. Long-term potentiation of horizontal connections provides a mechanism to reorganize cortical motor maps. J. Neurophysiol. 1994, 71, 2543–2547. [Google Scholar] [CrossRef] [PubMed]
- Hess, G.; Aizenman, C.D.; Donoghue, J.P. Conditions for the induction of long-term potentiation in layer II/III horizontal connections of the rat motor cortex. J. Neurophysiol. 1996, 75, 1765–1778. [Google Scholar] [CrossRef] [PubMed]
- Jones, E. GABAergic neurons and their role in cortical plasticity in primates. Cereb. Cortex 1993, 3, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Castro-Alamancos, M.A.; Connors, B.W. Short-term synaptic enhancement and long-term potentiation in neocortex. Proc. Natl. Acad. Sci. USA 1996, 93, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Bella, R.; Lanza, G.; Cantone, M.; Giuffrida, S.; Puglisi, V.; Vinciguerra, L.; Pennisi, M.; Ricceri, R.; D’Agate, C.C.; Malaguarnera, G. Effect of a gluten-free diet on cortical excitability in adults with celiac disease. PLoS ONE 2015, 10, e0129218. [Google Scholar] [CrossRef]
- Lanza, G.; Bella, R.; Cantone, M.; Pennisi, G.; Ferri, R.; Pennisi, M. Cognitive Impairment and Celiac Disease: Is Transcranial Magnetic Stimulation a Trait d’Union between Gut and Brain? Int. J. Mol. Sci. 2018, 19, 2243. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, G.; Lanza, G.; Giuffrida, S.; Vinciguerra, L.; Puglisi, V.; Cantone, M.; Pennisi, M.; D’Agate, C.C.; Naso, P.; Aprile, G. Excitability of the motor cortex in de novo patients with celiac disease. PLoS ONE 2014, 9, e102790. [Google Scholar] [CrossRef]
- Bella, R.; Ferri, R.; Lanza, G.; Cantone, M.; Pennisi, M.; Puglisi, V.; Vinciguerra, L.; Spampinato, C.; Mazza, T.; Malaguarnera, G. TMS follow-up study in patients with vascular cognitive impairment-no dementia. Neurosci. Lett. 2013, 534, 155–159. [Google Scholar] [CrossRef]
- Cantone, M.; Bramanti, A.; Lanza, G.; Pennisi, M.; Bramanti, P.; Pennisi, G.; Bella, R. Cortical Plasticity in Depression: A Neurochemical Perspective From Transcranial Magnetic Stimulation. ASN Neuro 2017, 9, 1759091417711512. [Google Scholar] [CrossRef]
- Lanza, G.; Bramanti, P.; Cantone, M.; Pennisi, M.; Pennisi, G.; Bella, R. Vascular cognitive impairment through the looking glass of transcranial magnetic stimulation. Behav. Neurol. 2017, 2017, 1–16. [Google Scholar] [CrossRef]
- Rioult-Pedotti, M.-S.; Friedman, D.; Donoghue, J.P. Learning-induced LTP in neocortex. Science 2000, 290, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Pleger, B.; Tegenthoff, M.; Ragert, P.; Förster, A.F.; Dinse, H.R.; Schwenkreis, P.; Nicolas, V.; Maier, C. Sensorimotor returning in complex regional pain syndrome parallels pain reduction. Ann. Neurol. 2005, 57, 425–429. [Google Scholar] [CrossRef]
- Hayman, M.; Kam, P.C. Capsaicin: A review of its pharmacology and clinical applications. Curr. Anaesth. Crit. Care 2008, 19, 338–343. [Google Scholar] [CrossRef]
- Cahill, L.; McGaugh, J.L. Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci. 1998, 21, 294–299. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Control | Capsaicin |
|---|---|---|
| 1 | 36 | 62 |
| 2 | 49 | 54 |
| 3 | 58 | 60 |
| 4 | 62 | 49 |
| 5 | 44 | 52 |
| 6 | 64 | 53 |
| 7 | 45 | 40 |
| 8 | 53 | 42 |
| 9 | 43 | 56 |
| 10 | 59 | 64 |
| 11 | 57 | 47 |
| 12 | 42 | 53 |
| Averages | 51.82 | 52.67 |
| SD | 9.02 | 7.40 |
| Control | Capsaicin | |||||
|---|---|---|---|---|---|---|
| Participant | Baseline | Post-Application | Post-Motor Learning | Baseline | Post-Application | Post-Motor Learning |
| 1 | 0.90 | 0.80 | 0.95 | 0.86 | 0.77 | 0.75 |
| 2 | 0.88 | 0.88 | 0.91 | 0.74 | 0.96 | 0.95 |
| 3 | 0.77 | 0.76 | 0.87 | 0.78 | 0.85 | 0.89 |
| 4 | 0.78 | 0.70 | 0.90 | 0.92 | 0.75 | 0.92 |
| 5 | 0.93 | 0.89 | 0.87 | 0.87 | 0.78 | 0.84 |
| 6 | 0.90 | 0.80 | 0.95 | 0.96 | 0.93 | 0.89 |
| 7 | 0.67 | 0.78 | 0.74 | 0.88 | 0.81 | 0.88 |
| 8 | 0.90 | 0.86 | 0.90 | 0.93 | 0.95 | 0.76 |
| 9 | 0.84 | 0.83 | 0.72 | 0.92 | 0.87 | 0.79 |
| 10 | 0.85 | 0.97 | 0.97 | 0.87 | 0.90 | 0.96 |
| 11 | 0.82 | 0.94 | 0.91 | 0.88 | 0.85 | 0.89 |
| 12 | 0.82 | 0.88 | 0,80 | 0.96 | 0.93 | 0.88 |
| Averages | 0.84 | 0.84 | 0.88 | 0.88 | 0.86 | 0.87 |
| SD | 0.073 | 0.077 | 0.081 | 0.067 | 0.073 | 0.069 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dancey, E.; Yielder, P.; Murphy, B. The Interactive Effect of Tonic Pain and Motor Learning on Corticospinal Excitability. Brain Sci. 2019, 9, 63. https://doi.org/10.3390/brainsci9030063
Dancey E, Yielder P, Murphy B. The Interactive Effect of Tonic Pain and Motor Learning on Corticospinal Excitability. Brain Sciences. 2019; 9(3):63. https://doi.org/10.3390/brainsci9030063
Chicago/Turabian StyleDancey, Erin, Paul Yielder, and Bernadette Murphy. 2019. "The Interactive Effect of Tonic Pain and Motor Learning on Corticospinal Excitability" Brain Sciences 9, no. 3: 63. https://doi.org/10.3390/brainsci9030063
APA StyleDancey, E., Yielder, P., & Murphy, B. (2019). The Interactive Effect of Tonic Pain and Motor Learning on Corticospinal Excitability. Brain Sciences, 9(3), 63. https://doi.org/10.3390/brainsci9030063