INTRODUCTION
Canopy-forming algae play important roles in habitat provision and primary production in temperate coastal ecosystems worldwide (Littler & Murray, Reference Littler and Murray1974; Thompson et al., Reference Thompson, Crowe and Hawkins2002). They are, however, under threat from a variety of impacts at local and regional scales including eutrophication (Bonsdorff et al., Reference Bonsdorff, Blomqvist, Mattila and Norkko1997; Rönnberg & Bonsdorff, Reference Rönnberg, Bonsdorff, Kautsky and Snoeijs2004), siltation resulting from urbanization (Schiel et al., Reference Schiel, Wood, Dunmore and Taylor2006; Airoldi & Hawkins, Reference Airoldi and Hawkins2007), harvesting (Ang et al., Reference Ang, Sharp and Semple1996; Zemke-White & Masao, Reference Zemke-White and Masao1999; Ugarte et al., Reference Ugarte, Sharp and Moore2006), trampling (Povey & Keough, Reference Povey and Keough1991; Keough & Quinn, Reference Keough and Quinn1998; Schiel & Taylor, Reference Schiel and Taylor1999; Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009) and point source pollution (Bellgrove et al., Reference Bellgrove, Clayton and Quinn1997; Keser et al., Reference Keser, Swenarton and Foertch2005). Canopy species are being influenced by the overarching effects of climate change interacting with local impacts, including rising and stormier seas (Underwood, Reference Underwood1998, Reference Underwood1999; Hawkins et al., Reference Hawkins, Sugden, Mieszkowska, Moore, Poloczanska, Leaper, Herbert, Genner, Moschella, Thompson, Jenkins, Southward and Burrows2009) and more frequent extreme high temperatures (Thompson et al., Reference Thompson, Crowe and Hawkins2002; Keser et al., Reference Keser, Swenarton and Foertch2005; Wernberg et al., Reference Wernberg, Bennett, Babcock, de Bettignies, Cure, Depczynski, Dufois, Fromont, Fulton, Hovey, Harvey, Holmes, Kendrick, Radford, Santana-Garcon, Saunders, Smale, Thomsen, Tuckett, Tuya, Vanderklift and Wilson2016), perhaps exacerbated by greater grazing pressure (Menge & Olson, Reference Menge and Olson1990; Davies et al., Reference Davies, Johnson and Maggs2007). Depending on the frequency of recurrence, these perturbations could act as either pulse disturbances (e.g. unusual spike in grazing pressure; Cervin & Åberg, Reference Cervin and Åberg1997), or long-term press disturbance (e.g. rising seas). Here we focus on the community consequences of a pulse disturbance that reduced the density of fronds (simulating breakage by herbivory or storm activity) of Ascophyllum nodosum (hereafter referred to as Ascophyllum), the major long-lived, canopy-forming macroalga dominating intertidal shores of the Northern Atlantic (Stephenson & Stephenson, Reference Stephenson and Stephenson1972; Jenkins et al., Reference Jenkins, Moore, Burrows, Garbary, Hawkins, Ingólfsson, Sebens, Snelgrove, Wethey and Woodin2008). Observational (Davies et al., Reference Davies, Johnson and Maggs2007; Gollety et al., Reference Gollety, Thiébaut and Davoult2011) and modelling studies (Hawkins et al., Reference Hawkins, Sugden, Mieszkowska, Moore, Poloczanska, Leaper, Herbert, Genner, Moschella, Thompson, Jenkins, Southward and Burrows2009) suggest Ascophyllum is likely to be increasingly disturbed over the coming decades in response to climate change and local anthropogenic impacts.
Individual Ascophyllum plants proliferate vegetatively from long-lived holdfasts, and stands are thought to last for centuries, with individual fronds lasting tens of years, creating a perennial habitat for epiphytic and understorey organisms (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009). Ascophyllum communities recover slowly after experimental manipulation (Jenkins et al., Reference Jenkins, Norton and Hawkins2004; Ingólfsson & Hawkins, Reference Ingólfsson and Hawkins2008) or harvesting (Ang et al., Reference Ang, Sharp and Semple1996; Zemke-White & Masao, Reference Zemke-White and Masao1999; Ugarte et al., Reference Ugarte, Sharp and Moore2006), taking more than a decade to return to their original state (Ingólfsson & Hawkins, Reference Ingólfsson and Hawkins2008).
In contrast to previous studies that examined the impacts of complete canopy removal (Bertness & Leonard, Reference Bertness and Leonard1997; Petraitis & Dudgeon, Reference Petraitis and Dudgeon1999; Jenkins et al., Reference Jenkins, Hawkins and Norton1999a, Reference Jenkins, Hawkins and Nortonb, Reference Jenkins, Norton and Hawkinsc, Reference Jenkins, Norton and Hawkins2004), we were interested in the impacts of partial frond loss, as might result from a pulse disturbance such as overgrazing, or an extreme weather event (e.g. heat wave, storm). The impacts of pulse disturbances resulting in partial canopy loss have been investigated for the fucoids Hormosira banksii (Schiel & Lilley, Reference Schiel and Lilley2007) and Fucus gardneri (Speidel et al., Reference Speidel, Harley and Wohnam2001), though have not previously been undertaken on Ascophyllum canopies or fronds.
Much attention has been given to press disturbances such as increased temperature due to climate change (Keser et al., Reference Keser, Swenarton and Foertch2005; Mieszkowska et al., Reference Mieszkowska, Hawkins, Burrows and Kendall2007), increases in grazing pressure (Pavia & Toth, Reference Pavia and Toth2000; Davies et al., Reference Davies, Johnson and Maggs2007) and trampling (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009), but a single pulse disturbance in full canopies can have long-term consequences at a local scale (Ingólfsson & Hawkins, Reference Ingólfsson and Hawkins2008; Wernberg et al., Reference Wernberg, Bennett, Babcock, de Bettignies, Cure, Depczynski, Dufois, Fromont, Fulton, Hovey, Harvey, Holmes, Kendrick, Radford, Santana-Garcon, Saunders, Smale, Thomsen, Tuckett, Tuya, Vanderklift and Wilson2016). Whilst full canopies of Ascophyllum can play an autogenic ecosystem engineering (sensu Jones et al., Reference Jones, Lawton and Shachak1994) role, we were interested in assessing the conditions under which this role breaks down, or the threshold frond density beyond which Ascophyllum ceases to be an effective ecosystem engineer. Specifically we tested the following hypotheses: (1) Temperature and light on the substratum will increase in proportion to frond reduction; (2) Changes in community composition will be proportional to frond reduction. Alternatively, (3) there is a threshold density of intact fronds below which Ascophyllum ceases to modulate temperature and light, with community composition showing a similar threshold.
MATERIALS AND METHODS
Spatial variation in assemblages associated with Ascophyllum nodosum
Three sites in southern England were initially surveyed in summer 2006: Batten Bay (Turnchapel) within Plymouth Sound (50°21′27″N 4°07′38″W), and two sites near the township of Looe (50°20′41″N 4°27′08″W), hereafter referred to as ‘Looe’ and ‘Hannafore Point’. All sites consisted of gently sloping wave-sheltered sedimentary rock platforms dominated at the mid-shore level by extensive canopies of Ascophyllum, inter-dispersed with patches of Fucus serratus and Fucus vesiculosus.
To assess the spatial variation in assemblages associated with a full canopy of Ascophyllum, all three sites were surveyed. Thirty replicate, haphazardly placed quadrats (0.3 × 0.3 m, appropriate to the scale of the macrofauna) with >95% Ascophyllum canopy cover (naturally lower cover canopies did not occur), were quantitatively sampled at each site for percentage cover of algae and sessile animals (using 49 point-intercepts, see Jenkins et al., Reference Jenkins, Coleman, Hawkins, Burrows and Hartnoll2005) and the abundance of individual mobile macroinvertebrates (defined as those >5 mm length), which were counted directly. Where algae and sessile animals occurred in quadrats but not under point-intercepts, they were recorded as present to account for rare species. In contrast to previous work that only examined a limited suite of species (Boaden & Dring, Reference Boaden and Dring1980; Bertness & Leonard, Reference Bertness and Leonard1997; Jenkins et al., Reference Jenkins, Hawkins and Norton1999a, Reference Jenkins, Norton and Hawkinsc; Leonard, Reference Leonard1999; Petraitis & Dudgeon, Reference Petraitis and Dudgeon1999; Speidel et al., Reference Speidel, Harley and Wohnam2001) or took a functional group response (e.g. ‘understorey species’; Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009), a complete inventory of the macrofauna and understorey algae of Ascophyllum stands was made and whole community responses measured (see also Gollety et al., Reference Gollety, Thiébaut and Davoult2011). The percentage covers of algal taxa were recorded as two separate layers: the canopy layer, and the understorey layer, which was revealed by gently moving the canopy aside.
Effects of a pulse disturbance reducing densities of Ascophyllum fronds on the physical environment and associated assemblages
A manipulative field experiment was conducted to simultaneously test the hypotheses that: (1) temperature and light on the substratum will increase in proportion to frond reduction; (2) changes in community composition will be proportional to frond reduction; or alternatively, (3) there is a threshold density of intact fronds below which Ascophyllum ceases to modulate temperature and light, with community composition showing a similar threshold. Hannafore Point was selected as the best site for experimental manipulation as it had an extensive bed of Ascophyllum, allowing good separation between sample plots, and it was easily accessible during low tide. Twenty haphazardly placed, fixed 4-m2 plots were established within a reef area of ~2000 m2, and labelled with plastic tags fixed with stainless steel screws into bedrock. All plots initially had >95% Ascophyllum cover and five replicates (all replicates are at the plot level) were assigned randomly to three frond reduction treatments and one unmanipulated control. The Ascophyllum holdfast occurs as a perennial basal disc from which multiple fronds arise directly in a clump, and can regrow after disturbance (Ugarte et al., Reference Ugarte, Sharp and Moore2006). The fronds are often very long (≥80 cm at our sites) and can create different amounts of canopy cover during low tide depending on where they have fallen on the receding tide. Therefore to allow a consistent disturbance to be applied between plots (mimicking frond breakage e.g. due to herbivory or storm activity) the frond density near the holdfast (rather than the percentage canopy cover) was manipulated. In July 2006, fronds in each holdfast clump were trimmed and removed to create the following treatments: 0% intact fronds (all fronds were removed), 25% intact fronds (75% of fronds were removed), 50% intact fronds (50% of fronds were removed) and 100% intact fronds (a control with unmanipulated frond densities). It is important to emphasize that the treatments represent the densities of intact fronds relative to the original frond densities and do not necessarily correspond to a linear relationship with canopy percentage cover. For each removal treatment, the fronds were cut at the base with garden secateurs to no more than 10 cm above the rock surface, to reduce frond density whilst still allowing for vegetative regeneration. Although these short fronds remained in all treatments throughout the experiment, they did not grow above 10 cm in length and no macroinvertebrates were recorded (unpublished data) using it as habitat. Any fronds able to sweep into the treatment plots from outside plants were trimmed back to prevent edge effects.
EFFECTS ON PHYSICAL ENVIRONMENT: TEMPERATURE AND LIGHT
Temperature was recorded on the substratum (under any canopy algae) of each experimental plot using a standard waterproof aquarium thermometer to the nearest 0.1°C on all sampling visits. Underwater TidBit Stowaway temperature data loggers (Onset, Bourne, MA, USA) were attached by bolts to the rock in six plots (six loggers unevenly spread over four treatments, N = 1–2 due to limited availability) to establish minimum and maximum temperatures within canopy treatments; these were set to record every 10 min over a 3-week period (2.8.2006–23.8.2006, summer). Light was recorded in all plots at sampling visits 7, 21 and 36 days as Photosynthetically Active Radiation (PAR) under the canopy (or simply on the substratum in 0% intact-frond plots) using a LI-COR-250 (LI-COR, Lincoln, NE, USA) meter with a waterproof sensor. All sampling was undertaken during daylight hours at low tide (summer sampling predominantly around midday). Further physical measurements were not taken on other sampling dates due to logistical constraints.
EFFECTS ON ASSOCIATED ASSEMBLAGES: MOBILE AND SESSILE MACROINVERTEBRATES AND MACROALGAE
The macroinvertebrate and macroalgal assemblages in each treatment were sampled using two 0.3 × 0.3 m sub-quadrats placed in the centre of the plot (fixed position, marked with a screw; randomized positioning was not possible without introducing inconsistencies in substratum complexity). Abundances of macroalgae, sessile and mobile macroinvertebrates in experimental plots were sampled 4 days before the manipulation (half of the plots were sampled on 1 day, half the following day due to logistical constraints, 19–20.6.2006) and 7, 21, 36, 69 and 272 days thereafter. Data from sub-quadrats were averaged to give one value per plot (N = 5 per treatment) to avoid pseudoreplication.
Data analysis
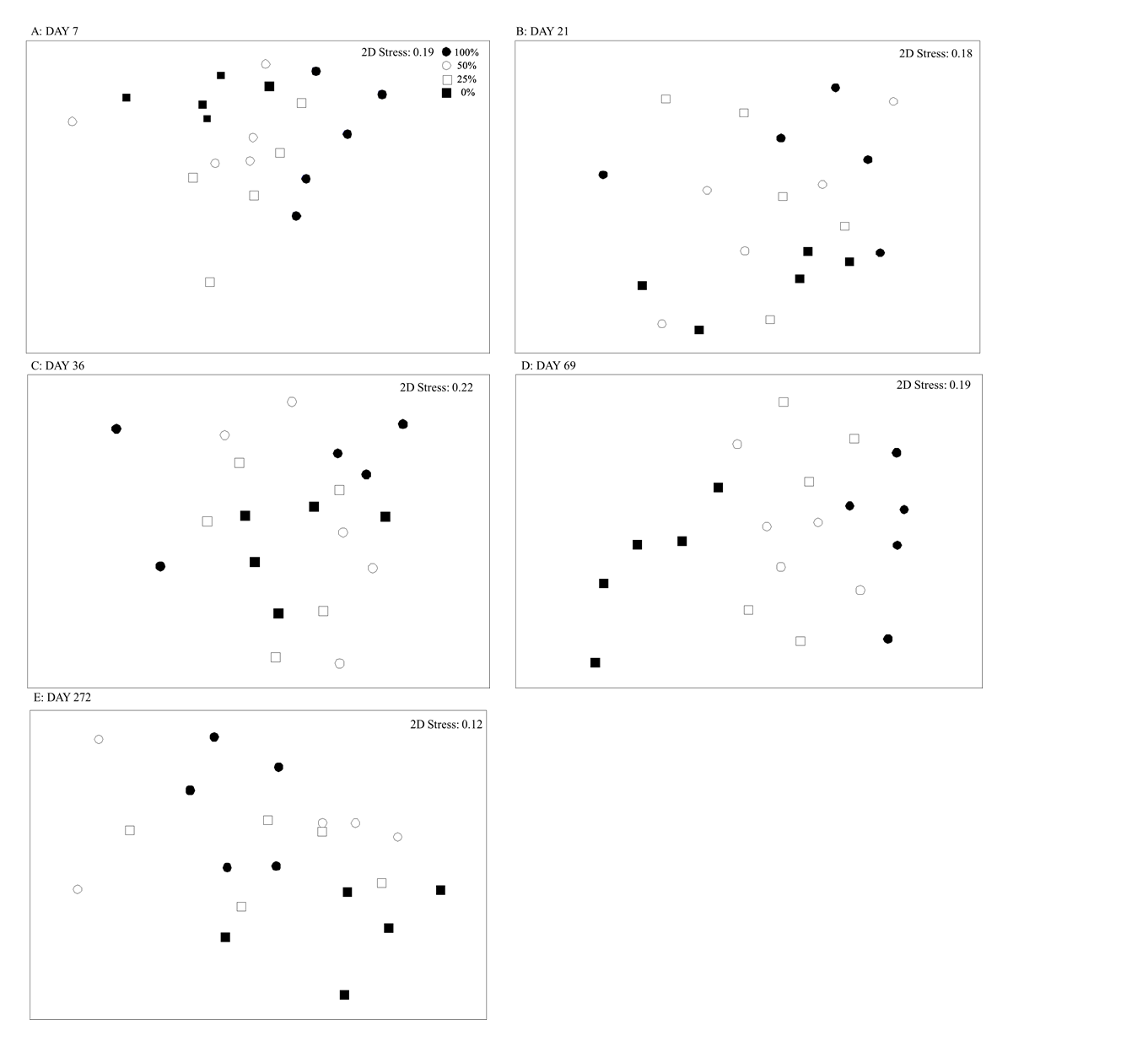
Multivariate analyses on species abundance and cover data (analysed separately) were used to compare (1) assemblages across sites (>95% Ascophyllum cover quadrats) at Hannafore Point (experiment site), Looe and Batten Bay; and (2) assemblages among experimental treatments at Hannafore Point. Differences between groups were visualized with nMDS based on Bray–Curtis similarities and ANOSIM identified the relative range of dissimilarities between groups (Bonferroni correction was applied; Quinn & Keough, Reference Quinn and Keough2002). SIMPER was applied to identify which taxa contributed most to the patterns identified in the ANOSIM and nMDS (contributions >10% are reported). All multivariate statistics were performed using Primer v7.
Univariate analyses comparing abundances of individual taxa in quadrats with naturally <95% canopy-cover of Ascophyllum between sites involved a single-factor ANOVA (Quinn & Keough, Reference Quinn and Keough2002) with Site as a random factor (Hannafore Point, Looe and Batten Bay). To determine which taxa and abiotic factors were affected by an experimental reduction of Ascophyllum fronds, differences among experimental treatments were analysed using a single-factor repeated-measures ANOVA (hereafter rmANOVA) (Quinn & Keough, Reference Quinn and Keough2002) with Treatment as a fixed factor. Response variables included species/taxon richness, individual species abundances, light and temperature. Where treatment × time interactions were significant, a one-way ANOVA and Tukey's test was undertaken for each sampling date to explain the differences between canopy controls and treatments (Quinn & Keough, Reference Quinn and Keough2002). All univariate tests were performed using SYSTAT v13.
Probability distribution plots were examined to visually test for normality of data, and residual plots were examined for homogeneity of variance (Quinn & Keough, Reference Quinn and Keough2002). Invertebrate count data were log (x + 1) transformed to better meet these assumptions. Algal covers were untransformed because, while they were proportions, the inclusion of primary and secondary cover mean that total cover was not limited to 100%, so the data did not follow a binomial distribution, for which a transformation would normally be used. As a check, analyses were done using arcsine-transformed data, and conclusions were identical. Untransformed data are presented here, because results are more easily interpretable. Where sphericity was not met in repeated-measures ANOVA, the Greenhouse–Geisser adjusted P-values were used (Quinn & Keough, Reference Quinn and Keough2002).
RESULTS
Spatial variation in assemblages associated with Ascophyllum nodosum
The assemblages of mobile invertebrates (ANOSIM, Global R = 0.155, P = 0.001) and sessile invertebrates and algae (Global R = 0.463, P = 0.001) differed amongst sites and all three sites were distinct (P = 0.001 for all pairwise comparisons). However, while statistically significant, the differences in the mobile invertebrate assemblages among sites were only slight (indicated by Global R and pairwise R-values <0.2). The periwinkles Littorina littorea and Littorina obtusata, and the topshell Phorcus lineatus differed amongst sites (ANOVA, respectively: F 2,87 = 15.991, P < 0.001; F 2,87 = 5.469, P = 0.006; F 2,87 = 11.310, P < 0.001). Littorina littorea accounted for 19–23% of the variation in mobile invertebrate assemblages among sites (mean ± SEM: 1.94 ± 0.12, 14.4 ± 0.35, 11.1 ± 0.40 per m2 at each of Hannafore Point, Looe and Batten Bay, respectively; SIMPER). Littorina obtusata accounted for 18–19% of the variation in mobile invertebrate assemblages among sites (mean ± SEM: 13.3 ± 0.30, 13.7 ± 0.28, 7.8 ± 0.21 per m2 at each of Hannafore Point, Looe and Batten Bay, respectively; SIMPER). Phorcus lineatus accounted for 11–12% of the variation in mobile invertebrate assemblages when comparing Hannafore Point and Looe with Batten Bay (mean ± SEM: 0.56 ± 0.12, 0.78 ± 0.10, 5.9 ± 0.25 per m2 at each of Hannafore Point, Looe and Batten Bay, respectively; SIMPER). Variability in the abundances of Patella spp., Gibbula cinerera and Actinia equina accounted for a further 13–19% of the variation in mobile invertebrate assemblages (SIMPER) but there were no differences in their mean abundances among sites (ANOVA, respectively: F 2,87 = 0.329, P = 0.720; F 2,87 = 0.841, P = 0.435; F 2,87 = 0.602, P = 0.550).
The differences in sessile invertebrate and algal assemblages among sites were stronger than those for mobile invertebrates (indicated by Global R and pairwise R-values between 0.3–0.6 for sessile species). Univariate analyses confirmed differences in the abundances of these species among sites, with Hannafore Point having the highest cover of Spirorbis spirorbis (ANOVA: F 2,87 = 17.3, P < 0.001, mean 2.7% ± 0.75) compared with Looe (mean 2% ± 0.70) and Batten Bay (mean 0.5% ± 0.30). Osmundea pinnatifida, encrusting coralline algae and bare rock were less abundant at Hannafore Point (mean 0.1% ± 0.12, <0.1% ± 0.03 and <0.25% ± 0.12, respectively) compared with both Looe (mean 2.7% ± 0.39, 4.8% ± 0.42 and 2.8% ± 0.45, respectively) and Batten Bay (mean 1.3% ± 0.37, 3.8% ± 0.46 and 2.3% ± 0.53, respectively: ANOVA; F 2,87 = 16.4, P < 0.001). Other sessile taxa accounted for less than 10% of the cumulative variation among sites (SIMPER) and were not investigated further.
Effects of a pulse disturbance reducing densities of Ascophyllum fronds on the physical environment and associated assemblages
EFFECTS ON PHYSICAL ENVIRONMENT: TEMPERATURE AND LIGHT
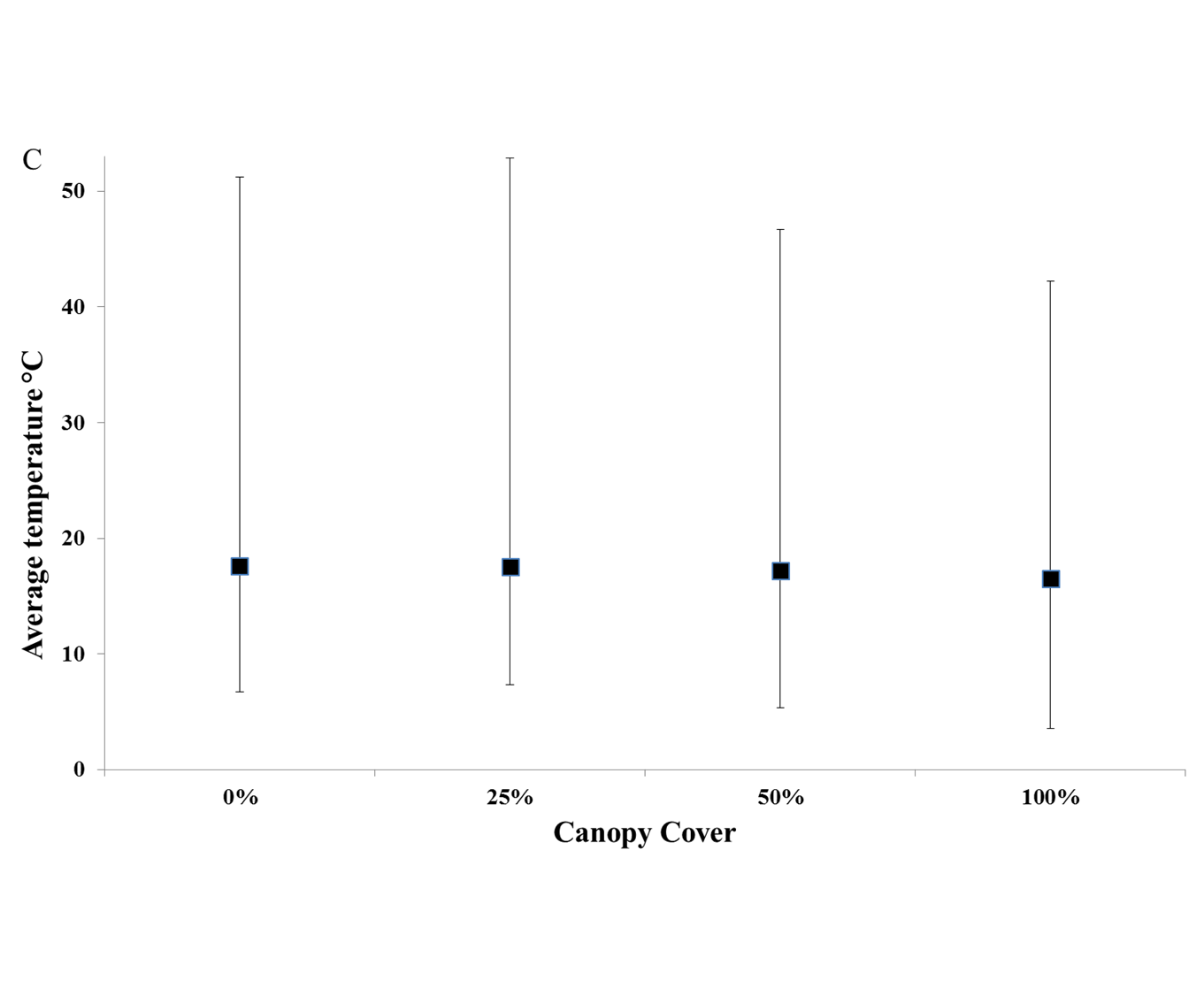
Temperatures recorded in the 50% intact-frond treatment did not differ from those found in control 100% intact-frond plots (mean 20°C ± 0.7 and 19°C ±0.5, respectively, Table 1: P > 0.05, Figure 1). The 25% intact-frond treatment had increased mean temperatures, at least 1°C above that recorded in the 100% intact-frond control (Table 1: P < 0.05, Figure 1). The 0 and 25% intact-frond treatments did not differ from each other with respect to temperature (P > 0.05) but both had increased mean temperatures, at least 1°C above that recorded in the 100% intact-frond control. The differences in average temperatures among treatments appear of a smaller magnitude in autumn and winter, but this was not supported statistically (time×treatment term, Table 1, Figure 1). The variation in temperatures among treatments was consistent over time with the maximum daily temperatures recorded in plots over a 3-week period (maximum, minimum and average temperatures are illustrated in Appendix C; no analysis possible due to low replication). Maximum daily temperatures recorded over this same period were lowest in 50 and 100% intact-frond treatments, and highest in both the 0 and 25% intact-frond treatments (Appendix C).

Fig. 1. Average temperature ± SE recorded on substratum beneath canopy layer (where present) within plots through time, N = 5. ● 100% intact-fronds, ○ 50% intact-fronds, □ 25% intact-fronds, ■ 0% intact-fronds.
Table 1. Repeated measures ANOVA comparing changes following disturbance to Ascophyllum canopy at Hannafore Point for time, treatment and time × treatment, N = 5. All df are: Treatment = 3(16), Time = 4, Treatment × Time = 12(64). MSResidual and degrees of freedom are provided to allow reconstruction of the full ANOVA table.

Statistical significance is indicated in bold, * indicates P-values adjusted to Greenhouse–Geisser Epsilon.
a Indicates significant Tukey's test (pairwise details described in Results text). Invertebrate count data were log (x + 1) transformed to meet assumptions of normality where necessary.
Available light (as PAR) in plots only increased after complete frond removal (0% intact fronds) when compared with the 100% intact-fronds control (Table 1, Figure 2). A trend towards increased light in the 25% intact-fronds treatment (Figure 2) relative to 100% intact-fronds control was evident when examining the means, but was not statistically significant (P > 0.05). Consistent with the patterns of variability in temperature among treatments, there was greater variability in available light amongst low frond-density treatments (0 and 25% intact fronds) compared with higher frond-density treatments (50 and 100% intact fronds) that had consistently low light levels (Figure 2).

Fig. 2. Average photosynthetically active radiation (PAR) ± SE recorded on substratum beneath canopy layer (where present) within plots through time, N = 5. ● 100% intact-fronds, ○ 50% intact-fronds, □ 25% intact-fronds, ■ 0% intact-fronds.
EFFECTS OF ASSOCIATED ASSEMBLAGES: MOBILE AND SESSILE MACROINVERTEBRATES AND MACROALGAE
Manipulation of the density of Ascophyllum fronds resulted in variable levels of canopy cover at low tide over time and the degree of disturbance to the fronds did not directly correspond with percentage cover (Table 1, Figure 3). The 50 and 100% intact-fronds treatments had similar Ascophyllum cover throughout the experiment (Table 1, Figure 3). From 7 to 36 days post-thinning, the 50% and 100% intact-fronds treatments had an average 85% Ascophyllum cover (Figure 3). Sixty-nine days post-thinning Ascophyllum declined to an average 77% cover in both treatments. The 25% intact-fronds treatment had lower Ascophyllum cover than both the 50% and 100% intact-fronds treatments, except at day 272 when a decline of Ascophyllum in the control and 50% intact-fronds treatment resulted in similar percentage cover (Table 1, Figure 3). The 25% intact-fronds treatment had much higher (mean ~60% throughout) Ascophyllum cover than the 0% intact-fronds treatment which had the lowest Ascophyllum cover (0%) of all treatments (Table 1, Figure 3).

Fig. 3. Average percentage cover ± SE of Ascophyllum canopy through time (not including holdfast fronds <10 cm), N = 5. ● 100% intact-fronds, ○ 50% intact-fronds, □ 25% intact-fronds, ■ 0% intact-fronds.
The assemblages of mobile and sessile taxa found in all plots were similar prior to experimental reductions of Ascophyllum (ANOSIM; Global R < 0, P > 0.6). Between 21 and 69 days after manipulation, assemblages of mobile or sessile taxa in the 50% intact-fronds treatment did not differ from those in the 100% intact-fronds control (Tables 2 & 3, Appendices A, B, D & E). Up to 70 days post-removal, the 25% and 0% intact-fronds treatments had different assemblages of mobile and sessile taxa compared with controls (Tables 2 & 3, Appendices A, B, D & E). On the final sampling visit (Day 272, ~9 months after the pulse disturbance, 16 April 2007, spring) the mobile invertebrate species assemblages were not different among treatments (Table 3). This reflected partial recovery of Ascophyllum in the 0% intact-fronds treatments (to ~25% canopy cover, Figure 3), and an increase in the percentage cover of the mid-canopy species Fucus serratus across all plots (~50% canopy cover, Figure 5C). There were, however, still significant differences after 9 months in the assemblages of sessile taxa in the 0% intact-fronds treatment compared with both the 25% and 100% intact-fronds treatments but the assemblages of sessile taxa in the 50% intact-fronds treatment did not differ from any other treatments (Appendix A).
Table 2. Pairwise ANOSIM comparing assemblages of sessile taxa (as percentage cover) among treatments (average for 2 subplots in each treatment area) at each sampling date, N = 5. Statistical significance (indicated by *) adjustment of alpha (Bonferonni correction) to 0.008, relative difference using R is indicated in bold.

Table 3. Pairwise ANOSIM comparing assemblages of mobile invertebrate species (invertebrate assemblage) among treatments (average for 2 subplots in each treatment area) at each sampling date, N = 5. Statistical significance (indicated by*) adjustment of alpha (Bonferonni correction) to 0.008, relative difference using R is indicated in bold.

All of the 10 mobile invertebrate species recorded (for full list see Appendix D) were found to contribute to differences identified in the ANOSIM analysis between the treatment types, although average abundances among treatment types and at different visits varied (further explained by univariate analyses below). Of all the sessile taxa (and/or cover components) sampled, Fucus serratus, encrusting corallines, bare rock, Spirorbis spirorbis, Osmundea pinnatifida, sand-covered rock, Mastocarpus stellatus, Ulva spp., Cladophora rupestris, Lomentaria articulata, unidentified red turf and the barnacle Perforatus perforatus, were found to contribute to the differences identified between canopy treatments (SIMPER). These individual species and habitat types were analysed using univariate techniques (when they occurred in sufficient abundances) to further identify responses to reduction of Ascophyllum (Table 1, Figures 4 & 5).

Fig. 4. Average abundances of invertebrates ± SE found in treatment plots through time (per m2), N = 5: (A) species richness; (B) Littorina obtusata; (C) Littorina littorea; (D) Phorcus lineatus; (E) Gibbula umbilicalis. ● 100% intact-fronds, ○ 50% intact-fronds, □ 25% intact-fronds, ■ 0% intact-fronds.
Invertebrate species richness in the 50% and 100% intact-fronds treatments were similar on days 7 and 21, with higher average species richness than both the 25% and 0% intact-fronds treatments (Table 1, Figure 4A, Tukey's P < 0.05). On day 36 only the 50% intact-fronds treatment differed from other treatments, with higher species richness (Table 1, Figure 4A, Tukey's P < 0.05). On day 69, the 100%, 50% and 25% intact-fronds treatments had similar average species richness, all higher than the 0% intact-fronds treatment (Table 1, Figure 4A, Tukey's P < 0.05). At the end of the experiment, the 0% intact-fronds treatment had lower species richness than the 50% and 100% intact-fronds treatments (Table 1, Figure 4A, Tukey's P < 0.05), but did not differ from the 25% intact-fronds treatment. The 25% intact-fronds treatment had similar species richness to the 50% intact-fronds treatment, but remained lower than the 100% intact-frond control (Table 1, Figure 4A, Tukey's P < 0.05).
SPECIES-SPECIFIC RESPONSES TO DISTURBANCE
The abundance of the periwinkle Littorina obtusata differed among treatments throughout the study (Table 1, Figure 4B). On day 7 the 25% and 0% intact-fronds treatments had less than half the abundance (~1/0.09 m2 compared with ~4/0.09 m2) of L. obtusata found in the 50% intact treatments and less than a quarter of that found in the 100% intact-fronds control (~10/0.09 m2, Figure 4B, Tukey's P < 0.05). On days 21 and 69, the 50% and 100% intact-fronds treatments had similar abundances of L. obtusata, which were higher than those recorded in both the 0% and 25% intact-fronds treatments (Figure 4B, Tukey's P < 0.05). The same pattern was evident on day 36, although not statistically significant (Table 1, Figure 4B). Littorina obtusata was more commonly observed on the fronds and holdfasts of Ascophyllum than under the canopy on the rock surface. In contrast, the abundance of the congeneric, largely rock-surface-dwelling, Littorina littorea did not respond to frond reductions but varied over time (Table 1, Figure 4C). The top shell Phorcus lineatus differed in abundance among treatments at different times throughout the study (Table 1, Figure 4D). The abundance of P. lineatus in the 0% intact-fronds treatment was higher than in the 100% intact-fronds control on days 21 and 36 and higher than that in all other treatments on day 69 (Figure 4D, Tukey's P < 0.05). The abundance of the top shell Gibbula umbilicalis varied among treatments on different sampling visits (Table 1, Figure 4E). On days 7 and 21 there were fewer G. umbilicalis in the 0% intact-fronds treatment compared with other treatments; although no differences were identified on other sampling visits (Table 1, Figure 4E, Tukey's P < 0.05).
Encrusting coralline algae did not differ (P = 0.066) in cover among treatments, though varied temporally, with a decline occurring on days 69 and 272 in all treatments (Table 1, Figure 5A). Although not quantified, a decline of the integrity of the encrusting coralline algae was observed in the 0% intact-fronds treatment, with a loss of pigment and surface peeling. Similarly, the cover of the turf-forming alga Osmundea pinnatifida did not vary among treatments, but varied over time, increasing in abundance on day 272 in the 100%, 50% and 25% intact-fronds treatments (Table 1, Figure 5B). The differences in abundance of the fucoid alga, Fucus serratus, among treatments and the control were not consistent over time (Table 1, Figure 5C). Fucus serratus had a higher percentage cover in the 100% intact-fronds control during day 7 compared with all other treatments; but no difference in percentage cover among treatments on other days (Table 1, Figure 5C, Tukey's P < 0.05). On day 272 a decline in abundance of F. serratus occurred in the control and 25% intact-fronds treatment and an increase occurred in the 0% intact-frond treatment (Table 1, Figure 5C). The percentage cover of the algae Ulva spp. only differed among treatments and the control plots on day 69 (Table 1, Figure 5D), with greater cover in the 100% intact-fronds control (average 33%) compared with the 50 and 25% intact-fronds treatments (both average 16%, Tukey's P < 0.05), Ulva spp. were not recorded in the control plots. Ulva spp. remained below 5% cover throughout the rest of the study. Univariate analyses of additional taxa found during the study were not possible due to low abundances.

Fig. 5. Average percentage cover of associated algae ± SE found in treatment plots through time, N = 5: (A) encrusting coralline algae; (B) Osmondea pinnatifida; (C) Fucus serratus; (D) Ulva spp. ● 100% intact-fronds, ○ 50% intact-fronds, □ 25% intact-fronds, ■ 0% intact-fronds.
DISCUSSION
Theory predicts that the importance or role of a species as an ecosystem engineer will depend on interactions and thresholds between environmental stress and a suite of morphological and ecological traits (Jones et al., Reference Jones, Lawton and Shachak1994, Reference Jones, Lawton and Shachak1997; Bruno & Bertness, Reference Bruno, Bertness, Bertness, Gaines and Hay2001; Crain & Bertness, Reference Crain and Bertness2006). Thus whilst a species may be considered to be an autogenic ecosystem engineer under one set of conditions (e.g. full canopy cover), it may cease to do so if these conditions break down. Intertidal rocky shores are particularly stressful environments with tidal cycles of emersion and gradients in physiological stress (e.g. desiccation, temperature, UV). Whilst full canopies of Ascophyllum have been demonstrated to act as autogenic ecosystem engineers and facilitate understorey biodiversity (Jenkins et al., Reference Jenkins, Hawkins and Norton1999a, Reference Jenkins, Hawkins and Nortonb, Reference Jenkins, Norton and Hawkinsc, Reference Jenkins, Norton and Hawkins2004; Jenkins & Hawkins, Reference Jenkins and Hawkins2003), we have demonstrated that this engineering role can be compromised by a pulse disturbance that reduces frond density and the ability of the canopy to modulate the physical environment. For Ascophyllum to act effectively as an autogenic ecosystem engineer at Hannafore Point, we found that at least 50% of the fronds (corresponding with 70–98% Ascophyllum cover) need to remain in order for temperature to remain cooler and for the associated invertebrate community to persist. With only 25% intact-fronds remaining (corresponding with 50–70% Ascophyllum cover), macroalgae and sessile invertebrates could persist at Hannafore Point but at reduced abundances.
Ascophyllum altered the physical environment by reducing light and temperature within the understorey, but the effect varied amongst pulse disturbance treatments and was not proportional to frond densities. Ascophyllum reduced temperature in the 50% and 100% intact-fronds treatments. The maximum temperatures recorded using the continuous data loggers also suggest that the canopy may buffer from temperature extremes since the 50% and 100% intact-fronds treatments had the least variation in temperature. This more stable temperature is likely to exert less physical stress on species associating with the canopy and improve survival (Crisp, Reference Crisp1964). Ascophyllum also filtered light to the understorey in all plots except those in which fronds were completely removed (0% intact-fronds treatment). Even the 25% intact-fronds treatment had an average light intensity six times lower than in the 0% intact-fronds treatment. Unexpectedly, percentage cover of Ascophyllum was not directly proportional to the density of fronds, and declined in all treatments with intact fronds over time. Throughout the experiment the percentage cover of Ascophyllum in the 50% and 100% intact-fronds treatments were similarly high, suggesting that Ascophyllum can withstand high levels of frond breakage whilst still maintaining a high canopy percentage cover and its ecosystem engineering function. This pattern tracked an ongoing decline in canopy cover in 100% intact-frond controls, suggesting additional natural influences on canopy cover occurring at this site, which may influence ecosystem engineering function.
A marked short-term disturbance effect was observed 7 days after the treatments were applied with species assemblages varying between all treatments and the control. Changes in species assemblages related to stable temperatures in controls compared with increased temperatures in all treatments. Light intensity to the understorey increased as fronds were reduced, although the 50% and 25% intact-fronds treatments had similar light intensities, and the 0% intact-fronds treatment had the highest light intensity (more than double the highest recorded in the 25–50% treatments). This short-term response for all data suggests the assemblage needs somewhere between 7 and 21 days to recover from a single disturbance. This contrasts with the findings of Speidel et al. (Reference Speidel, Harley and Wohnam2001) in Washington where abundances of littorinids, limpets and ephemeral algae did not differ among canopy reductions of Fucus gardneri despite rock temperatures being reduced by up to 7°C in all plots containing canopy algae. The differences in responses amongst these studies (Speidel et al., Reference Speidel, Harley and Wohnam2001 and this study) suggest that either particular species in Washington and the UK respond differently to canopy presence and/or rock temperature, or that rock temperature is not the driver of macroinvertebrate species patterns.
Our investigation found general similarities in responses of the community to those in Ascophyllum complete canopy-removal studies undertaken by Jenkins et al. on the Isle of Man (Jenkins et al., Reference Jenkins, Hawkins and Norton1999a, Reference Jenkins, Hawkins and Nortonb, Reference Jenkins, Norton and Hawkinsc, Reference Jenkins, Norton and Hawkins2004). In addition, we found that taxon richness was higher in the 100% and 50% intact-fronds treatments when compared with the 25% and 0% intact-fronds treatments. This pattern may also be related to differences in temperature amongst treatments. The algal turfs (e.g. Osmundea pinnatifida, Chondrus crispus) in the understorey of removed canopies on the Isle of Man (Jenkins et al., Reference Jenkins, Hawkins and Norton1999b) were found to degrade once the canopy was lost. We also found this at Hannafore Point (Lomentaria sp. and Cladophora sp.), but the species differed. Ingólfsson & Hawkins (Reference Ingólfsson and Hawkins2008) also found Cladophora sp. was lost following Ascophyllum removal in Iceland, and this loss persisted for 17 years despite canopy recovery. These studies in multiple locations suggest that a decline of turf-forming algae is a common response to Ascophyllum canopy loss and is likely a response to increased light and temperature when the canopy is removed (this study).
Although differences in abundance of Littorina obtusata occurred immediately after removal, subsequently abundances did not differ between the 50% and 100% intact-fronds treatments, but L. obtusata was much less common in the 25% and 0% intact-fronds treatments. Littorina obtusata occurs on the Ascophyllum canopy (Watson & Norton, Reference Watson and Norton1987; Williams, Reference Williams1992) and even displays a facilitative relationship with its ‘host’, as its grazing of tissue causes the alga to produce phlorotannins, reducing its susceptibility to grazing by other invertebrates (Pavia & Toth, Reference Pavia and Toth2000). Observations at all sites sampled in this study found a high number of L. obtusata attached to the fronds of Ascophyllum and since thinning and removal directly reduced fronds, this may explain lower abundances in the 25% and 0% intact-fronds treatments, especially at the beginning of the experiment.
Many of the taxa with reduced abundances in the 0% intact-fronds treatment were algae, suggesting that either higher light levels (Wiencke & Davenport, Reference Wiencke and Davenport1988; see Kubler & Raven, Reference Kubler and Raven1994; Irving et al., Reference Irving, Connell and Elsdon2004), or increased herbivory by grazers such as Phorcus lineatus (Mieszkowska et al., Reference Mieszkowska, Hawkins, Burrows and Kendall2007), impacted their survival in areas where the density of fronds was greatly reduced. De Figueiredo et al. (Reference De O. Figueiredo, Kain and Norton2000) reported that crustose corallines are common under algal canopies on rocky shores and in our study encrusting corallines bleached and peeled away from the surface in the 0% intact-fronds treatment, suggesting they are sensitive to high light, drying, temperature exposure or a combination.
Disturbances that may result in thinning of Ascophyllum canopy include increased grazing intensity (species introductions or range shifts, Davies et al., Reference Davies, Johnson and Maggs2007), climate change (reduced growth and recruitment) and trampling and harvesting (Boaden & Dring, Reference Boaden and Dring1980; Keser et al., Reference Keser, Swenarton and Foertch2005; Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009; Hawkins et al., Reference Hawkins, Sugden, Mieszkowska, Moore, Poloczanska, Leaper, Herbert, Genner, Moschella, Thompson, Jenkins, Southward and Burrows2009). Climate change is likely to affect Ascophyllum (Davies et al., Reference Davies, Johnson and Maggs2007) and shore communities in a multitude of ways such as increased sea and air temperatures stressing its physiological processes (Svensson et al., Reference Svensson, Pavia and Åberg2009), increased storm activity causing physical damage, and the immigration and emigration of species that may compete or alter species interactions as local conditions change (e.g. increased grazer diversity and abundance, Hawkins et al., Reference Hawkins, Sugden, Mieszkowska, Moore, Poloczanska, Leaper, Herbert, Genner, Moschella, Thompson, Jenkins, Southward and Burrows2009). The only published studies to examine thinning of Ascophyllum canopies were undertaken in Portugal and examined press disturbances by human trampling (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009, Reference Araújo, Sousa-Pinto, Serrão and Åberg2012; Bertocci et al., Reference Bertocci, Araújo, Vaselli and Sousa-Pinto2011). Araújo et al. (Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009) found trampling (at disturbance levels resulting in similar Ascophyllum cover to our 25% intact-fronds treatment) negatively impacted the community associated with Ascophyllum. Consistent with these studies (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009, Reference Araújo, Sousa-Pinto, Serrão and Åberg2012; Bertocci et al., Reference Bertocci, Araújo, Vaselli and Sousa-Pinto2011), we found a decreased abundance of some understorey species, but did not find increased Ulva spp. recruiting into 0% intact-fronds plots as seen in Portugal (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009). In contrast, Ulva spp. were in low abundance throughout the study at Hannafore Point with the exception of September 2006 (summer) where abundance spiked in all treatments except the 0% intact-fronds treatment. This is likely to be due to differences between sites and times of disturbance. Thinning of perennial intertidal fucoid algae was also investigated in New Zealand by Schiel & Lilley (Reference Schiel and Lilley2007) on the Australasian species Hormosira banksii. Schiel & Lilley (Reference Schiel and Lilley2007) found significant stepwise responses when comparing 0%, 25% and 50% canopy cover treatments to the 75% canopy and unmanipulated control plots. This matches with our findings at Hannafore Point where the macroinvertebrate assemblages found in our 100% and 50% intact-fronds treatments (which correspond, in terms of Ascophyllum canopy cover, with their 0% reduction) differed from those found in the 25% and 0% intact-fronds treatments (which correspond, in terms of Ascophyllum canopy cover, with their 75% and 100% reductions). Given the similarities between our findings and other studies, management (e.g. restricted site access for sensitive locations) may be necessary to maintain the ecosystem engineering function of Ascophyllum, especially as frond breakage due to multiple climate change impacts (e.g. increased desiccation/grazing pressure/storms) is predicted to increase (Hawkins et al., Reference Hawkins, Sugden, Mieszkowska, Moore, Poloczanska, Leaper, Herbert, Genner, Moschella, Thompson, Jenkins, Southward and Burrows2009).
Despite the spatial variation in species assemblages from undisturbed canopies detected between our study sites, other thinning studies with spatially variable assemblages have shown similar general patterns within the understorey including the loss of sensitive algae (e.g. non-geniculate corallines, Lilley & Schiel, Reference Lilley and Schiel2006), a decline in species richness (Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009) and replacement with other species (Jenkins et al., Reference Jenkins, Norton and Hawkins1999c; Araújo et al., Reference Araújo, Vaselli, Almeida, Serrao and Sousa-Pinto2009).
The results of this study support the general findings of other studies investigating the consequence of Ascophyllum canopy disturbance to shore ecology (Jenkins et al., Reference Jenkins, Hawkins and Norton1999a, Reference Jenkins, Hawkins and Nortonb, Reference Jenkins, Norton and Hawkinsc, Reference Jenkins, Norton and Hawkins2004; Jenkins & Hawkins, Reference Jenkins and Hawkins2003; Ingólfsson & Hawkins, Reference Ingólfsson and Hawkins2008), but additionally demonstrate that there is a threshold of disturbance beyond which the autogenic engineering role of Ascophyllum is compromised. Reduction of more than 50% of fronds near the holdfast will reduce its ability to modulate temperature and light and to support the understorey community assemblage. Given that intact Ascophyllum canopies have been shown to support high levels of biodiversity elsewhere (Black & Miller, Reference Black and Miller1991; Jenkins et al., Reference Jenkins, Hawkins and Norton1999b, Reference Jenkins, Norton and Hawkins2004; Keser et al., Reference Keser, Swenarton and Foertch2005; Davies et al., Reference Davies, Johnson and Maggs2008; Ingólfsson & Hawkins, Reference Ingólfsson and Hawkins2008; Watt & Scrosati, Reference Watt and Scrosati2013), disturbance resulting in frond loss is likely to have widespread consequences, with slow recovery times when less than 50% of fronds remain intact. Our work demonstrates the value of investigating the conditions under which habitat-forming species may modulate the physical environment and act as autogenic ecosystem engineers to better manage the assemblages and biodiversity that they facilitate.
SUPPLEMENTARY MATERIAL
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315416002009
ACKNOWLEDGEMENTS
Thanks are given to several field assistants particularly Kate de la Haye and technical assistants from The Marine Biological Association of the United Kingdom. In kind support was provided by The Marine Biological Association of the United Kingdom. Feedback from two anonymous reviewers improved this manuscript.
FINANCIAL SUPPORT
Funding was provided by the Ecological Society of Australia (student research award to J.B. Pocklington), Melbourne University (PORES and Drummond awards to J.B. Pocklington) and Museum Victoria (NHT grant to T.D. O'Hara). Additional funds were provided by The Marine Biological Association of the United Kingdom.